
Aust J Crop Sci. 18(09):547-554 (2024)
ISSN:1835-2707
https://doi.org/10.21475/ajcs.24.18.09.p77
Pattern of flower and fruit abscision and fruit growth of hass avocado under tropical conditions
Alexander Rebolledo Roa*, Ronal Arturo Burbano Díaz, Clara Inés Medina Cano
Corporación Colombiana de Investigación Agropecuaria-Agrosavia, Zip code 763533, Palmira, Valle del cauca, Colombia
ORCID: 0009-0006-6484-6699
Abstract
Research elucidating abscission processes in fruits of Hass avocados has been conducted in production areas in the subtropics, but there are few studies under tropical climate conditions that can serve as a starting point to guide suitable management practices. In a seven-year-old commercial orchard of Hass avocados, grafted onto native rootstocks (Antillean rootstocks), 20 trees were evaluated. A weather station installed on-site monitored the climate of the location. The pattern of fruit growth and development, flower and fruit abscission, and yield variables were characterized. More than 145,000 and 270,000 flowers abscised per tree were recorded, with final fruit sets of 0.1 ± 0.009% and 0.19 ± 0.04% in the first and second crop years, respectively. Fruit growth followed a sigmoid curve with no differences in growing degree-days until the time of harvest. Additionally, the alternate bearing behavior of Hass avocado was confirmed with an average yield of 24.3 ± 4.5 kg/tree and 93.3 ± 9.5 kg/tree for the first and second crop years, respectively, finding an average alternate bearing index of 0.74 ± 0.06.
Keywords: climate, early fruit drop, fruit development, fruit set, Persea americana.
Abbreviations: ABI_alternate bearing index, CEC_cation exchange capacity, EC_electrical conductivity, ET0_reference evapotranspiration. GDD_growing degree-days, PPT_ precipitation, SR_solar radiation, TEMP_temperature, WS_wind speed
Introduction
The area of avocado harvested worldwide in 2021 was 858,152 hectares with a production of 8,685,672 tons. For this year Colombia recorded an area of 94,111 hectares and a production of 979,617 tons, ranking as the second-largest avocado-producing country in the world (FAO Food and Agriculture Organization of the United Nations, 2021). According to the Köppen-Geiger classification, the climatic zones of the world are divided into five major groups, placing the tropical (megathermal) climate as Group A (Beck et al., 2018), where the avocado-producing areas in Colombia are located with temperatures above 18 °C and the annual precipitation exceeds evaporation (IDEAM, 2010).
The intensity of flowering in Hass avocado is initially affected by climatic factors such as frost, low or high temperatures, or water deficit stress as factors that determine the alternating bearing pattern of this species (Lovatt, 2010; Acosta-Rangel et al., 2021). In an on-crop year, there is marked flowering, a high percentage of fruit set, and a substantial yield, whereas in an off-crop year, there is limited flowering, a low percentage of fruit set, and a reduced yield (Mickelbart et al., 2012; Ziv et al., 2014; Garner and Lovatt, 2016).
The pattern of flower and fruit abscission and the factors influencing it has been extensively documented under subtropical conditions for Hass avocado (Lahav and Zamet, 1999; Garner and Lovatt, 2016; Alcaraz and Hormaza, 2021). An avocado tree can bear over a million flowers, with a fruit set rate between 0.01 and 0.1 for the Hass variety (Can-Alonzo et al., 2005; Garner and Lovatt, 2008; Alcaraz and Hormaza, 2014). The amount of fruit that ultimately remains on the tree until harvest depends on the endogenous nutritional and hormonal balance that is established in the plant between production cycles, the prevailing soil conditions, and the climatic and genetic factors (Garner and Lovatt, 2008; 2016).
Avocado fruit growth has been described as following a sigmoid curve (Dixon, 2007; Selladurai and Awachare, 2020). The development of avocados involves a sequence of stages: the initial phase, which may extend for about 10 weeks after full bloom; the exponential phase, which typically lasts around 30 weeks, contingent on the variety and environmental factors; and then the ripening phase, during which growth decelerates (Cowan et al., 2001). In California, Hass avocados have been reported to take 12 months to complete their fruit development (Garner and Lovatt, 2008) while in the Mexican states of Nayarit and Michoacán, this process usually lasts eight to nine months (Cossio-Vargas et al., 2008).
The main objective of this study was to generate information on the abscission of flowers and fruits, and the accumulation of fruit dry matter required to determine the best management strategies in avocado orchards to overcome it. The results reported here contribute to elucidating the production pattern and the alternate bearing behavior of Hass avocado under tropical conditions.
Results
Abscission and fruit growth
Flower abscission began in February and lasted until the beginning of June in both evaluated years. More than 145,000 flowers were recorded in the first year of evaluation and 270,000 in the second year. The abscission rate peaked in mid-April in both the first and second crop years (Fig. 3A and 3B). The number of flowers abscised per day was 2-fold lower in the first year compared to the second crop year. The abscised flowers represent more than 90% of the total reproductive structures in both years, with similar percentages between the two crop years (97.12 ± 0.49 and 97.38 ± 0.46, respectively). Fruit abscission began 40 days after flower abscission started in both crop years. The number of fruits abscised per day peaked in mid-May, with 170 ± 18 and 319 ± 50 in the first and second crop years, respectively (Fig. 3C and 3D). The pattern of immature fruit abscission was similar in both crop years, but the number of fruits abscised per day was 2-fold lower in the first crop year compared to the second crop year.
During both crop years, the flower abscission rate increased from February to April, where there was a predominantly negative water balance with PPT of 280 mm and 287 mm and ET0 of 311 mm and 302 mm in the first and second crop years, respectively (Figure 1E and 1F). Fruit abscission matched with an increase in PPT from May until August with 1623 mm and ET0 of 959 mm in the first-crop year and 1356 mm and 805 mm of PPT and ET0 in the second-crop year (Figure 1E and 1F).
In both crop years, fruit dry matter plotted against GDD resulted in a single sigmoid curve. The slow growth phase was recorded up to 300 GDD, with a weight ranging from 4.3 ± 0.13 to 5 ± 0.06 g. In the interval between 300 and 1600 GDD, the curve showed linear growth, with an increase in dry matter reaching weights between 47.2 ± 2.85 and 46.6 ± 1.57 g. After 1600 GDD, dry matter accumulation experienced a deceleration, revealing a more gradual increase until harvest, with weights ranging from 50.25 ± 0.35 to 50.38 ± 1.92 g. At the end of the growth cycle, it took 1789 GDD to reach harvest in the first crop year and 1900 GDD to reach harvest in the second crop year (Figure 2A and 2B).
The Boltzmann sigmoid model showed a very adequate fit for both crop years. The coefficients of determination (R2) exceeded 0.96 in both cases, and the correlations (r) exceeded 0.98 (Figure 2). These statistical results support the ability of the Boltzmann equation to accurately describe dry matter accumulation in fruit. Equations 4 and 5 characterize the behavior of dry matter accumulation during the two crop years, respectively (de Oliveira Aparecido et al., 2017).
$Fruit\ dry\ weigth\ (g) = \ - 1.153\frac{2.442 - ( - 1.153)}{1 + Exp\left( \frac{0.6877 - GDD}{0.6402} \right)}$ (4)
$Fruit\ dry\ weigth\ (g) = \ - 0.927\frac{2.002 - ( - 0.927)}{1 + Exp\left( \frac{0.6456 - GDD}{0.4538} \right)}$ (5)
The parameters calculated from the Boltzmann equation showed high significance at points a, b, c, d for both crop years, which corroborates the high R2 values presented by the model input data. In both cases, the parameters (Table 1) were similar and showed high significance, with values higher than P = 0.001 for all points. This suggests that the model could predict dry matter accumulation in avocado fruits with high accuracy and confidence under the conditions studied.
Harvest and alternate bearing index
All trees displayed alternate bearing in the two crop years, with a mean ABI of 0.74 ± 0.06. The harvest recorded in the first crop year was an average of 24.3 ± 4.5 kg/tree, and in the second crop year it was 93.3 ± 9.5 kg/tree. Despite the clear differences in production between crop years, a direct correlation could not be established between the harvest obtained in the first and second crop years. The percentage fruit set in the first crop year was 0.1% ± 0.009%, and in the second, the fruit set was 0.19% ± 0.04%. Figure 3 depicts the distribution of fruits at harvest according to the Codex Alimentarius standards for avocados. Forty-three (43)% of the fruits were of size 20 (134 - 147 g) for the first crop year while for the second crop year this percentage decreased to 35%. Furthermore, around 18% of the fruits corresponded to higher sizes between 28 and 30 for the first crop year while second crop year only 11% of the fruits were found within these sizes. As shown in Figure 3, in crop year 2 (represented by the dark bars), a higher percentage of fruits were in the lower size categories (18 - 24) compared to the fruits from crop year 1. The distribution of fruit weight at harvest showed a normal trend for both crop years (Shapiro-Wilk = 0.992, P = 0.051). In crop year 1, the recorded mean was 205 g ± 2.65, while in crop year 2, it was 184 g ± 2.45. The paired Student's t-test revealed highly significant differences between the two crop years (P < 0.001), with an average difference of 21 g ± 3.54.
Discussion
The climatic conditions of avocado-growing areas in Colombia show a small annual temperature range that is characteristic of tropical climates. Differences in average temperatures between the warmest and coldest months under tropical conditions rarely differ by more than 4 °C. The annual temperature range becomes greater at latitudes farther from the equator, but even at the limits of the tropical zone, few locations have an annual range greater than 15 °C (Trewin, 2014). The flower drop reported herein is possibly not related to heat stress because a daily maximum air temperature greater than 33 °C is required (Lomas, 1998; Garner and Lovatt, 2008).
On a seasonal time frame, central and western Colombia experience a double-peak annual precipitation pattern. There are distinct high rainfall periods during April-May and September-November, as well as low rainfall periods in December-February and June-August. These patterns are primarily influenced by the biannual movement of the intertropical convergence zone (ITCZ) (Poveda et al., 2011; Knoben et al., 2019). In the area of this study, low precipitation is confirmed from January to March and in June, in accordance with the reported bimodal behavior. This condition of low rainfall and high evapotranspiration could represent a stress factor during the flowering period under conditions of this study, since it has been reported in previous studies that avocado flowers include tepals with transpiratory activity under field conditions in southern California, which is attributed to the presence of functional stomata in the abaxial surface of the tepals (Blanke and Lovatt, 1993; Cárceles Rodríguez et al., 2023).
Under the conditions of this study, wind speeds of 25 km/h are present. According to the general classes of monthly wind speed data proposed by FAO (Allen, et al., 2006) the area is characterised by strong winds. This feature, in addition to creating conditions that favour a higher rate of transpiration, creates a higher drag force and may contribute to an increase in the fall of reproductive structures of the tree. (Gardiner et al. 2016). In addition, the reproductive success of Hass avocado is determined by pollination, factors such as nutrition, endogenous hormone content and stress factors also play crucial roles. In Colombian cultivation regions, a wide diversity of pollinating insects is reported, with those of the dipteran genus being prominent (Carabalí-Banguero et al., 2018). However, bees (Apis mellifera) appear to be the insects most involved in avocado flower pollination. A study conducted by installing beehives in field conditions showed that initial fruit set was significantly higher in crops with beehives, ranging between 0.048% and 0.058% (Peña and Carabalí, 2018), compared to the control which registered 0.028%. In our study, we observed a final fruit set of 0.1 ± 0.009% and 0.19 ± 0.04% in the first and second harvest years, respectively. These values are higher than those reported in the bee experiment. Importantly, our study did not employ beehives, but the high density of pollinating insects in Colombia appears to play a significant role in the findings (Carabalí-Banguero et al., 2018).
Flower and fruit abscission in this study lasted until June. In this period a mean value of 370 ± 17 W/m2 of SR was recorded with an increase of more than 100 W/m2 from June to September coinciding with the beginning of fruit formation process. In tropical environments, higher solar radiation is associated with lower cloud cover and higher temperatures (Ruiz et al., 2008) but in this study the temperatures do not exceed 25 °C. These conditions may promote carbohydrate availability for the developing fruit in accordance with previous studies indicating an increase in leaf net CO2 assimilation rate (Pn) above 20 mol·m-2·s-1 associated with cool days with temperatures below 30°C (Liu et al., 2002).
The soil's chemical properties in the study area, as described here, indicated a reduced Cation Exchange Capacity (CEC), resulting in limited cation availability. This situation is further influenced by the soil's acidic conditions. To meet the nutrient needs of the avocado trees, external nutrient applications were employed to supplement the nutrients removed during harvest and prevent excessive depletion of soil nutrient reserves (Rebolledo-Roa and Burbano-Diaz, 2023). This allowed us to avoid possible nutritional deficiencies as an additional stress factor for flower and fruit abscission, although previous studies in California reported no correlation with weather conditions or leaf nutrient concentration (Garner Lovatt, 2008, 2016).
Results obtained in this study showed a marked difference in the flowers number between crop years with a high flowering level in an on crop year and low flowering level in an off crop year, similar to other reports under subtropical conditions for Hass avocado (Garner and Lovatt, 2008). Salazar-García et al. (1998) reported that the high yield of trees during an "on" year significantly reduces the flowering intensity of the following productive cycle or "off" crop-year. The hypotheses that explain the effect of crop load on flowering in the following productive cycle are related to a greater depletion of carbohydrate reserves and an increase in the content of endogenous hormones, specifically gibberellins (Paz-Vega, 1997; Lovatt, 2006). This reduction was associated with a decrease in inflorescence production (13% of total buds/branch) which was accompanied by a high production of vegetative buds (72% of total buds/branch). Thus, vegetative growth fluxes occur at a lower magnitude in the "on" year, while in the "off" year vegetative growth is higher, this difference is considered to be a determinant of the alternate bearing behaviour in avocado (Salazar-García et al., 2006; Dixon, 2007).
Under the tropical climatic conditions of this study, the abscission timing found for Hass avocado was generally consistent with that reported for subtropical conditions in Hass and other cultivars with heavy flower and fruit abscission occurring during the spring and summer (Demirkol, 2002; Robinson et al., 2002; Alcaraz and Hormaza, 2021). A study conducted in a mesothermal climate in Araras, São Paulo State, Brazil, in Hass avocado also reported a specific timing of flower and fruit drop (Silva et al., 2017). The highest rate of small fruit drop reported in this study peaked in mid-May. Other studies have shown that the highest avocado fruit drop extends a few months after the end of the flowering season, followed by a gradually decreasing rate of fruit abscission that ceases in October (Garner and Lovatt, 2008; Silva et al., 2017). Our results agree with those reported by these studies, where fruit abscission continued until August in both crop years.
In the present study, the alternate bearing behavior of avocado was confirmed, with lower yield in the first crop year, which can be characterized as off-crop year, and higher yield in the second crop year, which can be characterized as on-crop year. Studies on alternate bearing in Hass avocado have been previously reported under subtropical conditions (Lovatt, 2010; Wolstenholme, 2010; Mickelbart et al., 2012; Sharma et al., 2019). The alternate bearing index reported in the present study coincides with that reported by Garner and Lovatt (2008), who found trees with a mean ABI of 0.72 ± 0.09. Typical ABI values for ‘Hass’ avocado orchards in California range between 0.57 and 0.97, which indicates that yield is 57% to 97% lower in the off-crop year than in the on-crop year (Lovatt, 2010). The alternate bearing index was quantified in Hass avocados grafted on ten clonal rootstocks (Borchard, D9, Duke 7, G755A, G755B, G755C, Topa Topa, Toro Canyon, G1033, Thomas), with the yield of trees on all rootstocks following a similar alternate bearing pattern but with values varying between 0.33 and 0.60 (Mickelbart et al., 2007). In our study, avocado trees were grafted on seedling rootstocks of the West Indian ecological group, with a possible influence on the increase in the ABI due to the high genetic variability of these rootstocks. In addition, this alternate bearing behavior may be disturbed by cultural practices such as postponement of avocado harvest or by local environmental events.
Differences in harvested fruit size have been reported in relation to crop load, termed on-crop years and off-crop years (Mickelbart et al., 2007; Lovatt, 2010; Garner and Lovatt, 2016). In the off-crop year, a 5% increase in fruit size ranging from 178 to 325 g was reported (Garner and Lovatt, 2008). In this study, an increase in larger fruit size for year one with low crop load was confirmed. In Hass avocado, competition for nutrients and carbohydrates between flowers and shoots as well as between fruits has been reported (Whiley, 1990; D’Asaro et al., 2021), with a higher availability of carbohydrate reserves in an off-crop year.
Materials and Methods
Field conditions
A total of twenty 'Hass' avocado trees, approximately six years old, were chosen from a commercial orchard located in the Rionegro municipality of the Antioquia department (06°05´56.9"N, 75°24´14.7"W) were selected based on qualitative characteristics such as high flowering level, vigor, and good phytosanitary conditions. The experimental area of this study is characterized by a tropical rainforest climate (Af), according to the Köppen-Geiger climate classification (Beck et al., 2018). Precipitation (PPT), maximum, mean and minimum temperatures (TEMP), solar radiation (SR), and daily wind speed (WS) were recorded with a weather station (Watch Dog 2900ET; Spectrum Technologies, Plainfield, IL, USA) and used to determine the daily reference evapotranspiration (ET0) based on the Food and Agriculture Organization (FAO) Penman–Monteith method and to calculate the water balance (Pereira, et al., 2015). The water balance followed a similar trend in both crop years, with a low-rain season and a negative water balance from January to April and June. The rest of the year had a positive water balance with the high-rain season periods occurring in May and from July to December (Figure 4). The crop years showed variations in average cumulative annual precipitation between the two years: 1988.61 mm in the first year and 1796.97 mm in the second year. The temperatures in the experimental orchard during both crop years exhibited a consistent pattern year-round, with average maximum, mean, and minimum temperatures of 23.7 ± 0.23 °C, 17.7 ± 0.22 °C, and 12.7 ± 0.16 °C, respectively (Figure 5A). The monthly average relative humidity was 71.8 ± 0.86%, and solar radiation showed a monthly average of 381.2 ± 17.0, with a high season from June to August and an annual average value of 445 ± 14.8 (Figure 5B).
The soils of the experimental area were mainly taxonomically classified as Typic Hapludan, by soils that are deep, well-drained, exhibit a texture ranging from moderately coarse to fine, and have a pH level that ranges from strongly to moderately acidic. These soils are characterized by their andic properties, originating from volcanic ash and igneous rocks (IGAC Instituto Geográfico Agustín Codazzi, 2009; Rebolledo-Roa and Burbano-Diaz, 2023). The chemical soil characteristics showed a moderately acidic pH (5.5 ± 0.02), high content of organic matter (OM: 19.8 ± 1.77%), and low cation exchange capacity (CEC) values (2.83 ±0.28 cmol(+)/kg). The macronutrients P, Ca, and Mg, as well as the micronutrients Mn and B, showed low concentrations, while the elements K, Cu, and Zn were found at normal levels. The electrical conductivity (EC) was 0.15 dS/m, indicating no salinity problems (Table 2).To determine the nutritional requirements of the experimental avocado orchard, the Kenworthy balance index method was used (Kenworthy, 1973; Rebolledo-Roa and Burbano-Diaz, 2023). Soil chemical analyses of the top 45 cm layer were conducted each year to assess the soil's nutrient supply capacity. Fertilization was carried out through soil application using easily assimilable sources of nutrients. A total of 682 g of N per plant was applied in the form of NH4 and urea, along with 262 g/plant of P2O5, 936 g/plant of K2O, 69.3 g/plant of CaO, and 193 g/plant of MgO. The orchard did not receive irrigation, and phytosanitary practices were performed by the farmer following integrated pest and disease management practices.
Abscission processes and fruit growth
Overall abscission was quantified in twenty trees with high flowering levels during two sequential crop years from full bloom in February until harvest in November. Black shade (at 70%) cloth was placed under each tree to cover the canopy area and every 8 days, the abscised flowers and developing fruits were hand-collected using brooms and dustpans. For each collection date, the total mass of the abscised structures was determined, three subsamples were taken and weighed, and the number of flowers and fruits was counted. The number of flowers and fruits for each subsample was used to determine its proportion with respect to the total mass on each collection date. The rate of flower and fruit abscission was determined by dividing the number of fruits and flowers collected on each collection date by the number of days between collection dates (Garner and Lovatt, 2008).
To determine the fruit dry weight, twenty inflorescences were randomly selected at stage 615, which corresponds to 50% of the flowers opened (Alcaraz et al., 2013), in the middle third of twenty trees with high flowering levels. The transverse diameter of the apical fruit in each inflorescence was recorded every 8 days with a digital caliper (MTC500-196, Mitutoyo Corp., Sakado, Japan) until it reached the dimensions specified for commercial maturity, as determined by the grower based on packing house standards (FAO, 1995; Rebolledo-Roa and Burbano-Diaz, 2023). The diameter was taken as a reference to collect twenty fruits according to the variability calculated from the respective standard error (SE) for each collection date. Fruits were packed in portable coolers before analysis, and the individual fruit fresh weight was determined using an analytical balance (Mettler AE200, USA). The fruit was dehydrated in a forced convection oven (Memmert UF 110, USA) at 70 °C until a consistent weight was achieved.
Harvest and alternate bearing index
The yield (kilograms/tree) and the quantity of fruits per tree were assessed for twenty trees at the time of harvest. The total number of fruits per tree was determined by weighing one hundred randomly selected fruits from trees with high crop loads. To express the degree of alternate bearing, the alternate bearing index (ABI) was calculated for the off- and on-crop years using the following Equation:
ABI = (yieldyear1− yieldyear2)/(yieldyear1+ yieldyear2) (1)
Statistical analysis
Climatic variables were examined using descriptive statistics and presented as monthly averages along with the standard error of the daily values for both crop years. The precipitation (PPT) and evapotranspiration (ET0) variables are shown as the monthly cumulative average over the two crop years. For soil analysis, means and SEs are reported for three samples. Fruit and flower abscission was analyzed using descriptive statistics and plotted as the average of each sampling date versus day of year. The fruit dry weight was plotted as the average of each sampling date versus cumulative growing degree-days (GDD). The GDD were estimated using Equation (2).
$GDD = \ \sum_{i = 1}^{n}{\ \ }(Tp - Tb)i$ (2)
GDD = Growing degree-days (°C)
i = Days
Tp = Daily average temperature
Tb = Base temperature at which growth stops (10 °C) (Aubert, 1972).
Additionally, some restrictions were considered:
If Tp > Tb ↠ GDD = Tp − Tb
For fruit dry weight, the Boltzmann model was used for fitting sigmoid data (De Oliveira Aparecido et al, 2017). This approach is particularly useful when the response variable is expected to gradually increase and then stabilize as the independent variable evolves over time (Farias and Dantas, 2022). Equation (3) is defined as follows:
$$\ y = \ d\frac{a - d}{1 + Exp\left( \frac{c - x}{b} \right)}\ (3)$$
where y: response variable (fruit dry weight), x: independent variable (GDD), a: upper limit of the response variable as x increases, d: lower limit of the response variable as x decreases, b: slope of the sigmoid curve, c: point on the curve where the rate of change of y is most pronounced (De Oliveira Aparecido et al, 2017).
To evaluate the significance of these four points, their behavior was compared between both crop years. This comparison was carried out to determine if significant changes in the trend curve and points were manifested.
The fruits harvested were categorized according to the Codex Alimentarius standards for avocados (FAO Food and Agriculture Organization of the United Nations, 1995). A paired Student's t-test was performed with a significance level of P = 0.05 to compare the crop years. Statistical analyses were performed using R-Project for Statistical Computing, version 4.2, and conducted within the RStudio environment, version 2022.07.1.
Conclusions
Under the conditions of this study adjusted to the tropical rainforest climate (Af) classification, the flower and fruit abscission patterns showed the same trend throughout each production cycle, as reported under subtropical conditions. The weather recorder showed a constant trend throughout the year that did not suggest an influence of environmental conditions on the pattern of flower and immature fruit abscission, since no extreme climatic conditions were recorded. The low rain season coinciding with the flowering phase at anthesis can act as a stress factor due to the increase in transpiration caused by the stomatal activity of the flower tepals. There was a clear difference in the flower and fruit set numbers between the two crop years, which allowed the identification of year one as an off-crop year and year two as an on-crop year. The crop load of the trees influences the harvest distribution pattern, with larger fruit size in the off-crop year. The fruit growth in dry weight fitted to a sigmoid curve as described under subtropical conditions.
Acknowledgments
The authors would like to thank Colciencias designated the Corporación Colombiana de Investigación Agropecuaria-Agrosavia as the executing public entity of the project “Integrated management of Hass avocado crops (Persea americana) to increase quality and yield in Colombian production areas".
References
Acosta-Rangel A, Li R, Mauk P, Santiago L, Lovatt CJ (2021) Effects of temperature, soil moisture and light intensity on the temporal pattern of floral gene expression and flowering of avocado buds (Persea americana cv Hass). Sci Hortic. 280: 109940.
Alcaraz ML, Hormaza JI (2021) Fruit set in avocado: Pollen limitation, pollen load size, and selective fruit abortion. Agronomy. 11(8): 1603.
Alcaraz ML, Hormaza, JI (2014) Optimization of controlled pollination in avocado (Persea americana Mill, Lauraceae). Sci Hortic. 180: 79–85.
Alcaraz ML, Thorp TG, Hormaza JI (2013) Phenological growth stages of avocado (Persea americana) according to the BBCH scale. Sci Hortic. 164: 434–439.
Allen RG, Pereira LS, Raes D, Smith M. (2006). Evapotranspiración del cultivo: guías para la determinación de los requerimientos de agua de los cultivos. FAO. 298.
Aubert B (1972) Considerations sur la phenologie des especes fruitieres arbustives. Fruits 27(3): 193–198.
Beck HE, Zimmermann N, McVicar TR, Vergopolan N, Berg A, Wood E F (2018) Present and future Köppen-Geiger climate classification maps at 1-km resolution. Scientific Data. 5(1): 180214.
Blanke M, Lovatt CJ (1993) Anatomy and Transpiration of the Avocado Inflorescence. Ann Bot. 71(6): 543–547.
Can-Alonzo C, Quezada-Euán JJ, Xiu-Ancona P, Moo-Valle H, Valdovinos-Nunez GR, Medina-Peralta S (2005) Pollination of ‘criollo’ avocados ( Persea americana) and the behaviour of associated bees in subtropical Mexico. J Apicult Res. 44(1): 3–8.
Carabalí-Banguero D , Montoya-Lerma J, Carabalí-Muñoz A. (2018) Dípteros asociados a la floración del aguacate Persea americana Mill cv. Hass en Cauca, Colombia. Biota Colombiana. 19(1): 92-111.
Cárceles Rodríguez B, Durán Zuazo VH, Franco Tarifa D, Cuadros Tavira S, Sacristan PC, García-Tejero IF (2023) Irrigation alternatives for avocado (Persea americana Mill) in the Mediterranean subtropical region in the context of climate change: A Review. Agriculture. 13(5): 1049.
Cossio-Vargas LE, Salazar-García S, González-Durán IJ, Medina-Torres R (2008) Fenología del aguacate “Hass” en el clima semicálido de Nayarit, México. Rev Chapingo Ser Hort. 14(3): 319–324.
Cowan AK, Cripps RF, Richings EW, Taylor NJ (2001) Fruit size: Towards an understanding of the metabolic control of fruit growth using avocado as a model system. Physiol Plantarum. 111(2): 127–136.
D’Asaro A, Reig C, Martínez-Fuentes A, Mesejo C, Farina V, Agustí M (2021) Hormonal and carbohydrate control of fruit set in avocado ‘Lamb Hass’ A question of the type of inflorescence?. Sci Hortic. 282: 110046.
De Oliveira Aparecido LE, Ferreira RV, Souza Rolim G, De Souza BS, De Souza PS (2017) Nonlinear agrometereological models for estimating lychee fruit growth. Rev Bras Frutic. 39(2).
Demirkol A (2002) Phenological and pomological characteristics and yields of some avocado cultivars grown in Antalya ecological conditions Anadolu. Ege Tarimsal Arastirma Enstitusu Mudurlugu, Aegean Agricultural Research Institute Directorate. (2002): 49-64.
Dixon J (2007) Shoot growth of ‘Hass’ avocado trees in ‘on’and ‘off’flowering years in the western bay of plenty NZ Avocado Growers Association Ann. Res Report. 7: 41–48.
Dixon J, Lamond CB, Smith DB, Elmlsy TA (2006) Patterns of fruit growth and fruit drop of ‘Hass’ avocado trees in the Western Bay of Plenty, New Zealand New Zealand Avocado Growers’ Assn Ann. Res Report. 6: 47–54.
FAO Food and Agriculture Organization of the United Nations (1995) Codex Alimentarius standard (Codex Stand 197, 1995).
FAO Food and Agriculture Organization of the united nations (2021) FAOSTAT Retrieved September 10, 2023, from https://wwwfaoorg/faostat/en/#home website: https://wwwfaoorg/faostat/en/#home.
Farias LA, Dantas BF (2022) Morphometric characterization and functional traits of fruits and seeds of Neoglaziovia variegata (Arruda) Mez. Journal of Seed Science. 44.
Gardiner B, Berry P, and Moulia B. (2016). Wind impacts on plant growth, mechanics and damage. Plant Sci. 245: 94-118.
Garner L, Lovatt CJ (2008) The relationship between flower and fruit abscission and alternate bearing of ‘Hass’ avocado. J Am Soc Hortic Sci. 133(1): 3–10.
Garner L, Lovatt CJ (2016) Physiological factors affecting flower and fruit abscission of ‘Hass’ avocado. Sci Hortic. 199: 32–40.
IDEAM (2010) Atalas Climatologico de Colombia 1981 - 2010 Retrieved September 10, 2023, from http://atlasideamgovco/visorAtlasClimatologicohtml website: http://atlasideamgovco/visorAtlasClimatologicohtml.
IGAC Instituto Geográfico Agustín Codazzi (2009) Estudio general de suelos y zonificación de tierras de Cauca Bogotá: IGAC.
Kenworthy, AL (1973) Leaf analysis as an aid in fertilizing orchards In Soil testing and plant analysis (pp 381–392) Madison: Soil Sci Soc Amer.
Knoben WJ, Woods RA, Freer JE (2019) Global bimodal precipitation seasonality: A systematic overview. Int J Climatol. 39(1): 558–567.
Lahav E, Zamet D (1999) Flowers, fruitlets and fruit drop in avocado trees. Rev Chapingo Ser Hort. 5: 95–100.
Liu X, Sievert J, Arpaia ML, Madore MA. (2002) Postulated physiological roles of the seven-carbon sugars, mannoheptulose, and perseitol in avocado. J Am Soc Hortic Sci. 127(1): 108-114.
Lomas J (1988) An agrometeorological model for assessing the effect of heat stress during the flowering and early fruit set on avocado yields. J Am Soc Hortic Sci. 113(1): 172–176.
Lovatt CJ (2010) Alternate bearing of ‘Hass’ avocado Calif Avocado Soc Yearb 93: 125–140.
Lovatt CJ. (2006) Eliminating alternate bearing of the ‘Hass’ avocado. Paper presented at proceedings of the california avocado research symposium. University of California, California 127-142.
Mickelbart MV, Bender GS, Witney GW, Adams C, Arpaia ML (2007) Effects of clonal rootstocks on ‘Hass’ avocado yield components, alternate bearing, and nutrition The. J Hortic Sci Biotech. 82(3): 460–466.
Mickelbart MV, Robinson PW, Witney G, Arpaia ML (2012) ‘Hass’ avocado tree growth on four rootstocks in California II Shoot and root growth. Sci Hortic. 143: 205–210.
Paz-Vega S. (1997) Alternate bearing in the avocado (Persea americana Mill.). California Avocado Society Yearbook. 81: 117-148.
Peña JF, Carabalí A. (2018) Effect of Honey Bee (L.) Density on pollination and fruit set of avocado (Mill.) cv. Hass. J Apic Sci. 62(1): 5-14.
Pereira LS, Allen RG, Smith M, Raes D (2015) Crop evapotranspiration estimation with FAO56: Past and future. Agr Water Manage. 147: 4–20.
Poveda G, Álvarez DM, Rueda Ó A (2011) Hydro-climatic variability over the Andes of Colombia associated with ENSO: a review of climatic processes and their impact on one of the Earth’s most important biodiversity hotspots. Clim Dynam. 36(11–12): 2233–2249.
Rebolledo-Roa A, Burbano-Diaz RA (2023) Nutrient Balance in Hass Avocado Trees as a Tool to Optimize Crop Fertilization Management. Agronomy. 13(8): 1956.
Robinson PW, Mickelbart MV, Liu X, Adams C, Witney G, Arpaia ML (2002) Development of a phenological model of avocado tree growth in California. Acta Hortic. 575: 859–864.
Ruiz D, Moreno HA, Gutiérrez ME, Zapata PA (2008) Changing climate and endangered high mountain ecosystems in Colombia. Sci Total Environ. 398(1-3), 122-132.
Salazar-García S, Cossio-Vargas LE, Lovatt CJ, González-Durán IJ, Pérez-Barraza MH (2006) Crop load affects vegetative growth flushes and shoot age influences irreversible commitment to flowering of Hass' avocado. Hort Science. 41(7): 1541-1546.
Salazar-García S, Lord E M, Lovatt CJ. (1998) Inflorescence and flower development of the ‘Hass’ avocado (Persea americana Mill.) during ‘on’ and ‘off’ crop years. J Am Soc Hortic Sci. 123(4):537-544.
Selladurai R, Awachare CM (2020) Nutrient management for avocado (Persea americana Miller). J Plant Nutr. 43(1): 138–147.
Sharma N, Singh SK, Mahato AK, Ravishankar H, Dubey AK, Singh NK (2019) Physiological and molecular basis of alternate bearing in perennial fruit crops. Sci Hortic. 243: 214–225.
Silva SR, Cantuarias-Avilés TE, Chiavelli B, Martins MA, Oliveira MS (2017) Phenological models for implementing management practices in rain-fed avocado orchards1. Pesquisa Agropecuária Tropical. 47(3): 321–327.
Trewin B (2014) The climates of the tropics and how they are changing. State of the Tropics 1: 39–52.
Whiley A (1990) Interpretación de la fenología y fisiología del palto para obtener mayores producciones Producción, Postcosecha y Comercialización de Paltas. Curso Internacional 1–25.
Wolstenholme BN (2010) Alternate bearing in avocado: an overview Pietermaritzburg, South Africa University of KwaZulu-Natal.
Ziv D, Zviran T, Zezak O, Samach A, Irihimovitch V (2014) Expression profiling of FLOWERING LOCUS T-Like Gene in alternate bearing ‘Hass’ avocado trees suggests a role for PaFT in avocado flower induction. Plos One. 9(10): e110613.
Table 1. Estimated parameters of the Boltzmann sigmoidal model for two curves of Hass avocado dry weight.
| Crop year 1 | Crop year 2 | ||||||
|---|---|---|---|---|---|---|---|
| Estimate | SE | t -value | Pr >|t| | Estimate | SE | t-value | Pr>|t| |
| a 2.442 | 0.12036 | 20.30 | <2e-16 | a 2.002 | 0.08390 | 23.86 | <2e-16 |
| b 0.640 | 0.04193 | 15.27 | <2e-16 | b 0.453 | 0.03140 | 14.45 | <2e-16 |
| c 0.687 | 0.04552 | 15.11 | <2e-16 | c 0.645 | 0.03344 | 19.30 | <2e-16 |
| d -1.153 | 0.04807 | -23.99 | <2e-16 | d -0.927 | 0.03112 | -29.80 | <2e-16 |
Table 2. Soil nutritional content in avocado orchards located in Rionegro (Antioquia - Colombia)
| Mean | SE | |
|---|---|---|
| pH | 5.5 | 0.02 |
| EC (ds/m) | 0.15 | 0.01 |
| OM (%) | 19.81 | 1.77 |
| P (mg/kg) | 4.3 | 0.70 |
| S (mg/kg) | 7.4 | 1.68 |
| Ca (cmol+/kg) | 1.71 | 0.18 |
| Mg (cmol+/kg) | 0.78 | 0.09 |
| K (cmol+/kg) | 0.28 | 0.02 |
| Na (mg/kg) | 0.07 | 0.00 |
| CEC (cmol+/kg) | 2.83 | 0.28 |
| B (mg/kg) | 0.18 | 0.01 |
| Cu (mg/kg) | 1.5 | 0.09 |
| Fe (mg/kg) | 300 | 10.35 |
| Mn (mg/kg) | 1.3 | 0.09 |
| Zn (mg/kg) | 3 | 0.30 |
Figure 1. Number of avocado flowers and immature fruits abscised per day. (A, C) Crop year 1. (B, D) Crop year 2. Error bars represent the SE for that collection period. (E) Water balance for crop year 1. (F) Water balance for crop year 2. ET0, reference evapotranspiration (FAO Penman‒Monteith method); PPT, monthly accumulated precipitation.
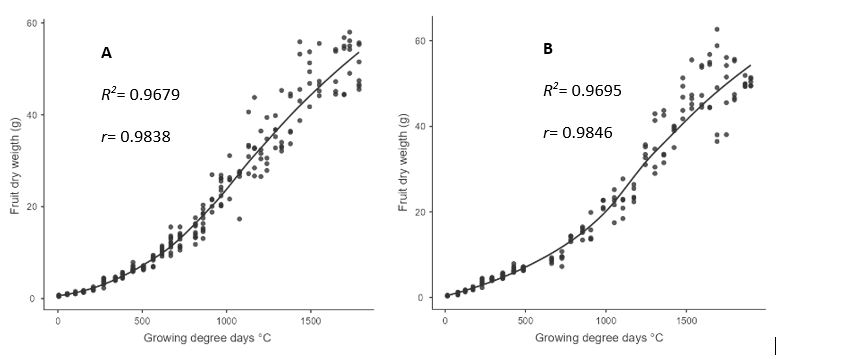
Figure 2. Fruit dry weight accumulation from anthesis to harvest and absolute growth rate for the first crop year (A) and second crop year (B).
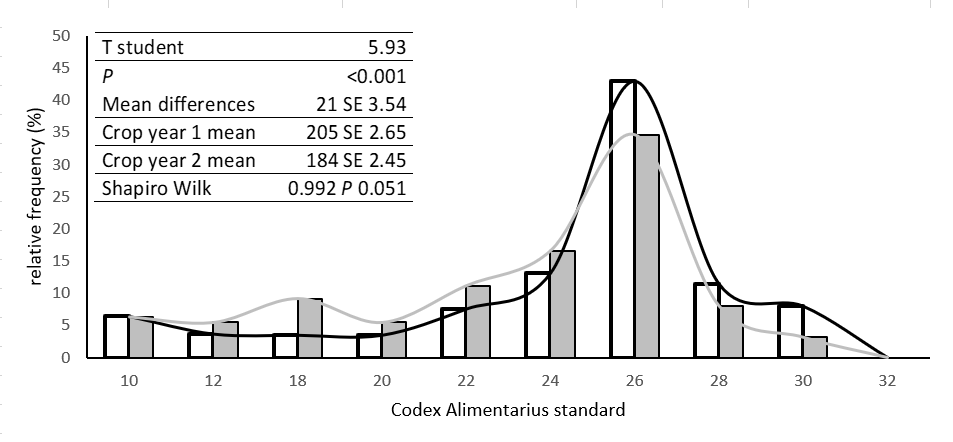
Figure 3. Classification of Hass avocado harvest, relative frequency of weights according to the Codex Alimentarius standard for avocado.
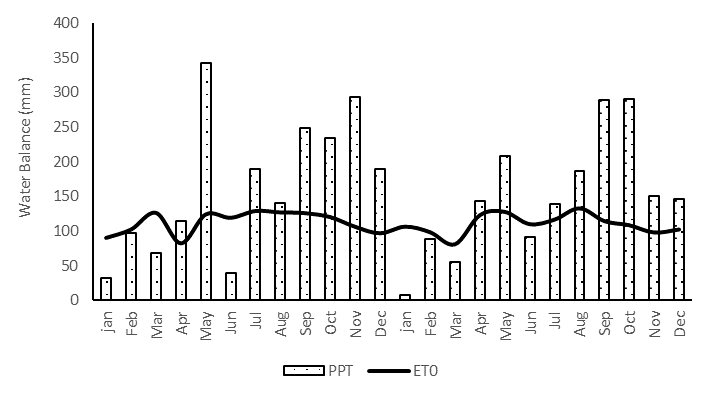
Figure 4. Water balance for Hass avocado experimental orchards in Rionegro (Antioquia). ET0: reference evapotranspiration, reference evapotranspiration (Pereira et al., 2015); PPT: precipitation.
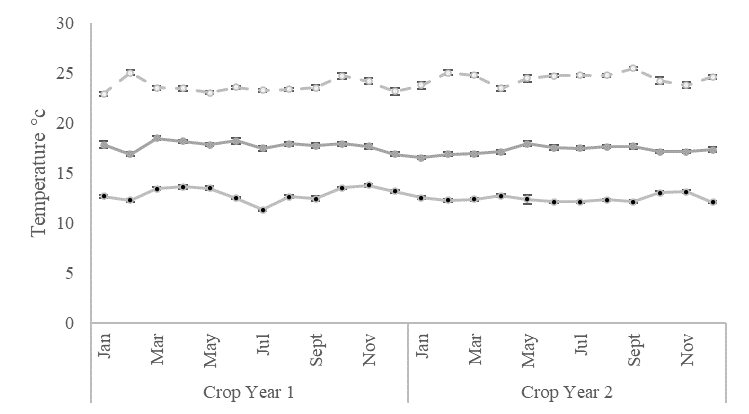
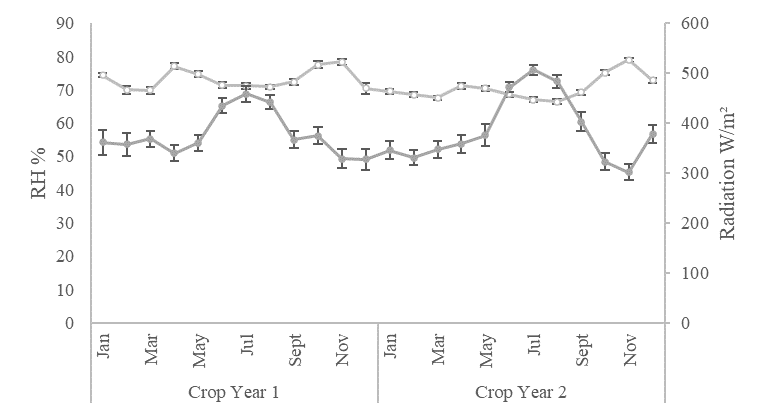
Figure 5. Climatic variables for Hass avocado experimental orchards in Rionegro (Antioquia). (A) Maximum, mean, and minimum temperatures. (B) Relative humidity and radiation.
