
Aust J Crop Sci. 18(11):731-739 (2024) | ISSN:1835-2707
https://doi.org/10.21475/ajcs.24.18.11.p106
Intra and interspecific competition in spatial arrangements of cassava and peanut intercropping
Maria Beatriz Bernardes Soares1, Everton Luis Finoto1, José de Anchieta Alves de Albuquerque2*, Glauber Ferreira Barreto2, João Luiz Lopes Monteiro Neto2, José Maria Arcanjo Alves2, Richard Alcides Molina Alvarez2, Laura Soliane Cruz Braz2, Anderson Carlos de Melo Gonçalves3, Edgley Soares da Silva4
1Agência Paulista de Tecnologia dos Agronegócios/ Polo Centro Norte, Pindorama, SP, Brasil
2Universidade Federal de Roraima/Centro de Ciências Agrárias, Boa Vista, RR, Brazil
3Instituto Federal de Educação, Ciência e Tecnologia/ Campus Coari, Coari, AM, Brasil
4Instituto de Educação e Inovação, Boa Vista, RR, Brazil
*Corresponding author: José de Anchieta
Alves de Albuquerque  | ORCID ID:
https://orcid.org/0000-0003-4391-258X
| ORCID ID:
https://orcid.org/0000-0003-4391-258X
Abstract: Intercropping cassava with peanuts in different plant arrangements can increase cassava crop yield. Nonetheless, no experimental evidence has been established on the ubiquitous forces of intra and interspecific competitions affecting cassava development, crop yield, and land use efficiency (LUE) in intercropping. Therefore, this study aimed to evaluate the intra- and interspecific competition impacts on cassava culture, crop productivity, and LUE under different spatial arrangements in the cassava-peanut consortium. The experiment was arranged in randomized blocks with 5 treatments and 5 replications. The treatments were: cassava in monoculture (T1); cassava + peanuts between the cassava rows (T2); cassava + peanuts in a double row between the cassava rows (T3); cassava planted in a double row + peanuts between double rows of cassava (T4); peanuts in monoculture (T5). The results reveal that intercropping affects the vegetative growth rate of cassava and that denser spatial arrangements promote greater inter- or intraspecific competition between cassava and peanut plants, affecting the height, branching, root length, diameter, and commercial yield of cassava roots. However, less dense arrangements provide more advantageous conditions for cassava root growth, resulting in less root loss. The productivity of cassava and peanut roots and grains was higher in single cultivation. The intercrop of cassava in a single row with peanuts in the alley and the intercrop of cassava in a double row with peanuts presented satisfactory ELU, with an average of 1.30.
Keywords: Arachis hypogaea L.; Intercropping; Land use efficient; Manihot esculenta Crantz.
Introduction
Grown mainly in tropical and subtropical regions, cassava (Manihot esculenta Crantz) is one of the most important sources of carbohydrates and dietary energy for human nutrition. It is also considered a resistant plant, as it can adapt and produce in edaphoclimatic conditions that limit the cultivation of more profitable crops (Rubiano-Rodríguez and Cordero-Cordero, 2019; Reichert et al., 2021).
Despite cassava's hardiness, production can be affected by its intrinsic growth characteristics, such as slow development in the initial phase, long harvesting cycle, yield variation between cultivars, and the crop spatial arrangement. These factors favor weed incidence competing for growth resources available in the environment (water, light, nutrients), leading to productivity losses of up to 100% when not controlled, as well as exposing the soil to erosive agents that accelerate erosion losses (Albuquerque et al., 2012; Lima et al., 2015; Legodi and Ogola, 2020; Reichert et al., 2021).
In these conditions, the intercropping system, also known as poly cropping or mixed cropping, is a sustainable agricultural practice that can reduce crop production losses because, among its many advantages, this system promotes better use of growth resources, less use of cultivated area and contributes to the reduction of weeds, insect pests, and diseases. These advantages convert into superior yield results compared to monoculture (Albuquerque et al., 2015; Legodi and Ogola, 2020; Amoako et al., 2022). The architecture of the vegetative canopy and the plant population directly influence crop growth and production (Oliveira et al., 2019).
Different intercropping models are suggested for cassava cultivation. Nonetheless, intercropping cassava with leguminous plants is more widespread due to the beneficial effects achieved, such as those reported by Alves et al. (2010), Albuquerque et al. (2012), Legodi & Ogola (2020) and Amoako et al. (2022). The use of legumes in intercropping systems has a wide range of benefits, mainly by improving the supply of soil nitrogen for accompanying crops, favoring the process of soil conservation, controlling weeds, maximizing area use, and increasing the producer profit (Alves et al., 2010; Albuquerque et al., 2012; Mudare et al., 2022).
It is possible to use several indicators to compare intercropping and monoculture systems. However, the index conventionally used is Land Use Efficiency (LUE), which evaluates the intercropping benefits and efficiency compared to monocultures (Agegnehu et al., 2006), considering the use of available resources and agricultural productivity.
It is noteworthy that intercropping success is not only influenced by the growth characteristics of the intercropped crops, their life expectancy, the period of coexistence, and the crops' spatial distribution combination but also by intra- and interspecific competition forces governing the plants' coexistence in the mixed crop (Albuquerque et al., 2012; Maitra et al., 2021; Legodi and Ogola, 2020). Due to these factors, studies suggest that the cassava and legumes intercropping in different plant arrangements differ in production and LUE (Dapaah et al., 2003; Albuquerque et al., 2012).
Nevertheless, there is a lack of concrete experimental evidence on the effect of intercropping cassava and peanuts on the development of cassava plants during the growing phase. The peanut (Arachis hypogaea L.) is a legume (fabaceous) of great importance in some regional cultivation systems, being the second most grown oilseed after soybeans, accounting for 9% of world oilseed production (Peoples et al., 2021). In Brazil, the state of São Paulo is the largest peanut producer (Botelho et al., 2020) and the third largest cassava producer (IBGE, 2022), indicating how crucial the cultivation of these crops represents for the economy and local and regional supply.
Therefore, understanding the performance of the cassava crop and the viability of intercropping with peanuts is essential to obtaining more accurate information about the production of these crops. This study aimed to assess the effects of intra- and interspecific competition between cassava and peanut spatial arrangements on cassava crop development, crop yields, and LUE.
Results and Discussion
The effects of intraspecific competition between cassava plants in dense cultivation and interspecific competition between cassava and peanut plants cultivated between lines can be seen in the growth rate of the cassava, especially in the first 90 days, where treatments T1 and T3 showed similar behavior, but with an average daily growth rate 11.28% higher than the treatments T2 and T4 (Fig 1). This behavior may be related to the lower strength of intra- and interspecific competitions between the cassava and peanut crops due to less dense spatial arrangement in these treatments (T1 and T3) (Fig 1). This growth rate pattern can be attributed to resource competition during the peanut grain filling phase and cassava root growth, especially water since this evaluation period happened between May and July when water availability in the soil was lower.
However, after 90 days from planting (DAP), in treatments T2 and T4, the cassava plants increased their growth rate, surpassing the plants in treatments T1 and T3 (Fig 1). Possibly, the cassava dense planting in the T2 treatment and T4 double-row cassava cultivation system, provided greater competition from this period onwards, which may have caused the etiolation of the cassava canopy, affecting the growth rate parameters (tx and Am.tx/e) and the time needed to reach the maximum daily growth rate (DAPm) (Table 1).
In the T2 and T4 growing arrangements, the tx and Am.tx/e values are, on average, 34.47% and 18.16% higher than the parameters in the T1 and T3 treatments. In addition, for the DAPm values, the T1 and T3 treatments take an average of 20 days longer to reach the maximum growth rate (Table 1). In general, for all the cultivation arrangements, the growth rate is highly affected in the third of the crop cycle (90 DAP) and in the maximum growth period, which in this study averaged 156 DAP (Fig 1), respectively, when cassava plants enter the stages of high light interception and maximum light interception (Leihner, 1983). Cassava grown in intercropping achieves approximately 40% of light interception in the first 90 DAP, with maximum light interception between 150 and 210 DAP, with a decrease afterward (Leihner, 1983). However, the cassava crop's initial, maximum, and decreasing periods of light interception can vary between cultivars depending on plant size, leaf angle, and growing season (Mahakosee et al., 2022).
Although the cultivation arrangements T2 and T4 provided higher values for the parameters tx and Am.tx/e, for the parameters Am and Am/e, the highest values were observed in the treatments T1 and T3, indicating that in these cultivation conditions, the cassava plants have higher potential for growth in height and a higher growth rate at the inflection point of the curve. These results partially support the final plant height values (Table 2).
The results of the cultivation arrangements on final plant height, stem diameter, and number of stems per plant are displayed in Table 2. As can be seen, the T1 arrangement resulted in a similar final plant height to the value observed in T3 and was higher than the values achieved in the T2 and T4 arrangements (p < 0.05). However, the final plant height in T3 was also similar to the value in the T2 arrangement (p < 0.05).
Higher values observed for final plant height indicate a possible relationship with the growth potential regarding the maximum value achieved for this variable (Table 1), which converges into final plant height, observed mainly in less dense intercropping (T1 and T3) for cassava (Table 2). Results disagreeing with the ones found in this study are reported by Amoako et al. (2022), who observed no differences in cassava plant height when intercropped with cowpea, regardless of the time the cassava plant height was sampled (90 and 180 DAP and during harvest). Therefore, depending on the species intercropped with cassava and the cultivation arrangement, there may be an influence on light sharing and, consequently, changes in plant development (Mahakosee et al., 2022).
Concerning stem diameter, only the arrangement where the cassava plants were cultivated at a reduced spacing between rows (T4) caused a significant decrease in this variable. The other treatments did not differ statistically from each other (p < 0.05), even though, on average, they provided plants with a larger diameter (3.0 cm) when compared to the T4 system (Table 2). However, this was not the case for the number of stems per plant, where the arrangements tested did not cause any significant differences (p < 0.05) (Table 2).
These results suggest that the stem diameter reduction inversely follows the plant's growth rate (tx and Am.tx/e). The cassava plants grown in the T4 cultivation system experienced a phase of rapid elongation of the main stem and principal sympodial branches, resulting in stem elongation caused by intraspecific competition. However, the rapid vertical growth of these structures did not lead to an increase in the number of total stems, the final plant height (Table 2), or the length of the secondary sympodial branches.
The lack of significance of the plant stem number may be related to the trichotomous branching type and establishment period of the IAC 576-70 cultivar, which practically stabilized 30 days before the evaluation period. The present stem diameter observations partially corroborate the findings of Mansaray et al. (2022) and are opposite to the plant stem number, as observed by Akpan & Ikeh (2018). The latter authors found that the number of stems on the cassava decreases depending on the cultivar, regardless of the spacing between cassava plants.
Regarding the branching data (Table 3), only treatment T2 stood out with the highest branching percentage, with an average 4.42% higher than the other treatments (T1, T3 and T4) (p < 0.05). The branching percentage is a characteristic
Table 1. Estimates of Gompertz equation parameters and their relationships for plant height in different spatial arrangements of cassava cultivation. Pindorama, SP.
| Spatial arrangement | Maximum potential height | Height at inflection point | Maximum relative rate | Maximum growth rate | Maximum growth day |
| Am (m) | Am/e (m) | tx(mm.m-1) | Am.tx/e (mm.day-1) | DAPm (day) | |
| T1 - Cassava monoculture | 3.03 | 1.11 | 12.95 | 14.44 | 171.31 |
| T2 - Cassava + Peanuts between lines | 2.53 | 0.93 | 19.56 | 18.21 | 148.96 |
| T3 - Cassava + Peanuts in double lines | 2.97 | 1.09 | 14.66 | 16.02 | 160.41 |
| T4 - Cassava + Peanuts in dense double lines | 2.29 | 0.84 | 22.56 | 19.01 | 143.39 |
Table 2. Final plant height, stem diameter, and stem number per plant of the cassava cultivar IAC 572-70 at 270 DAP under the influence of cultivation systems. Pindorama, SP.
| Cultivation systems | Final plant height (cm) | Stem diameter (cm) | Stem number per plant |
| T1 - Cassava monoculture | 255.96 a | 2.92 a | 2.44 a |
| T2 - Cassava + Peanuts between lines | 237.91 b | 3.14 a | 2.68 a |
| T3 - Cassava + Peanuts in double lines | 249.13 ab | 3.02 a | 2.68 a |
| T4 - Cassava + Peanuts in dense double lines | 223.41 c | 2.44 b | 2.24 a |
| F | 16.555** | 10.1219** | 0.7810ns |
| CV (%) | 5.96 | 7.49 | 14.70 |
1 Averages followed by the same letter do not differ significantly by Tukey's test (p≤0.05).
Table 3. Branching, and height of the first sympodial branch (RS1) and of the second sympodial branch (RS2) at 270 DAP of cassava cultivar IAC 572-70 under the influence of different cultivation systems. Pindorama, SP.
| Cultivation systems | Branching (%) | Height RS1 (cm) | Height RS2 (cm) |
| T1 - Cassava monoculture | 84.38 b | 39.91 a | 116.75 a |
| T2 - Cassava + Peanuts between lines | 88.62 a | 27.08 b | 90.58 b |
| T3 - Cassava + Peanuts in double lines | 85.98 b | 34.83 a | 97.91 b |
| T4 - Cassava + Peanuts in dense double lines | 83.74 b | 36.25 a | 112.25 a |
| F | 102.04** | 62.355** | 17.894** |
| CV (%) | 3.21 | 13.22 | 4.89 |
1Averages followed by the same letter do not differ significantly by Tukey's test (p≤0.05).
that matters since the branches are used as propagative material for future crops. However, the branching amount varies depending on the cultivar, the size of the plant, and the legume species in the consortium (Mansaray et al., 2022).
For TB1 height, treatment T2 showed the lowest observed value, with an average 26.80% lower than the other cultivation systems T1, T3 and T4 (p < 0.05). For SB2 height, treatments T1 and T4 showed similar values, though, with 17.68% higher than T2 and T3 (p < 0.05). However, these treatments (T2 and T3) did not differ from each other, with a variation of 90.58 to 97.91 cm height per plant in RS2 (p < 0.05) (Table 3).
The height variation in RS1 and RS2 reflects the plant's rate of leaf and branch generations during its growth, which influences the cassava crop development throughout the cultivation cycle (Fagundes et al., 2009; Gabriel et al., 2014). Streck et al. (2014) found that cassava plants grown in monoculture at different row spacings reduce the leaf number in denser spacing and increase it in wider ones, which may have influenced the partitioning of photoassimilates to meet the growth of certain plant parts to the detriment of others (Gabriel et al., 2014). Thus, as observed during the experiment, the cassava plants in the T2 and T4 cultivation systems had smaller leaves and lower SB2 height, although with a higher percentage of branching and secondary sympodial branches (Fig 2).
The cultivation systems had a significant effect (p < 0.05) on the main stem and primary and secondary sympodial branch lengths. The main stem was 26.75% shorter when the cassava plants were grown in the T2 cultivation system compared to the T1, T3 and T4 treatments. However, there was no significant difference (p < 0.05) between treatments T1, T3 and T4 (Fig 2). For the main sympodial branches, treatments
T1 and T4, and systems T2 and T3, provided similar average lengths when compared to each other (p < 0.05). Nonetheless, the main sympodial branches in the T2 and T3 treatments were 17.14% shorter when compared to the lengths of the main sympodial ones in the T1 and T4 cultivation systems (p < 0.05) (Fig 2).
Concerning the length of secondary sympodial branches (Fig 2), the T2 treatment stood out showing an average 17.95% higher than the other treatments (T1, T3 and T4) (p < 0.05). The higher percentage of branches (Table 3) in the T2 cultivation system was probably influenced by the secondary branches’ length since their elongation caused the plant’s self-competition by light radiation. Moreover, the plants presenting the tallest main stem also displayed the shortest secondary sympodial branches (Fig 2). Silva et al. (2017) support that the number of plant branches can change depending on the first insertion height. Besides, cassava plants allocate more photoassimilates to the roots when the main stem has fewer secondary branches, reducing the partitioning competition between root and shoot, which results in higher root productivity (Silva et al., 2017).
The cultivation systems had a significant influence (p < 0.05) on root length, root diameter, the percentage of non-commercial roots (Losses %), and the yield of the cassava crop (Table 3). Concerning root length, the cultivation configurations T2 and T3 show statistically equivalent averages (p < 0.05) (Table 4). However, on average, these roots are 22.72% longer than the average length of the ones produced in treatments T1 and T4, which did not differ statistically (p < 0.05).
Regarding the root diameter, the T3 cultivation system provided the highest average (5.05 cm) and was 8.11% higher than the T1 and T4 treatments and 17.82% higher than the T2 arrangement. Meanwhile, the T1 and T4 cultivation
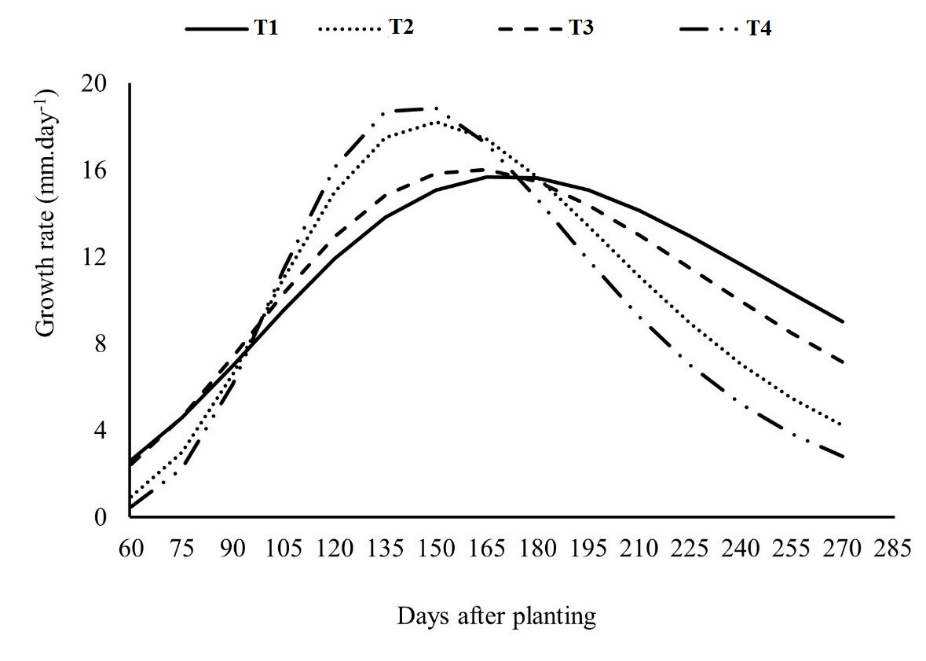
Fig 1. Growth rate of cassava plants under the influence of cultivation systems: monoculture (T1) or intercropping with peanuts (T2, T3 and T4). Pindorama, SP.
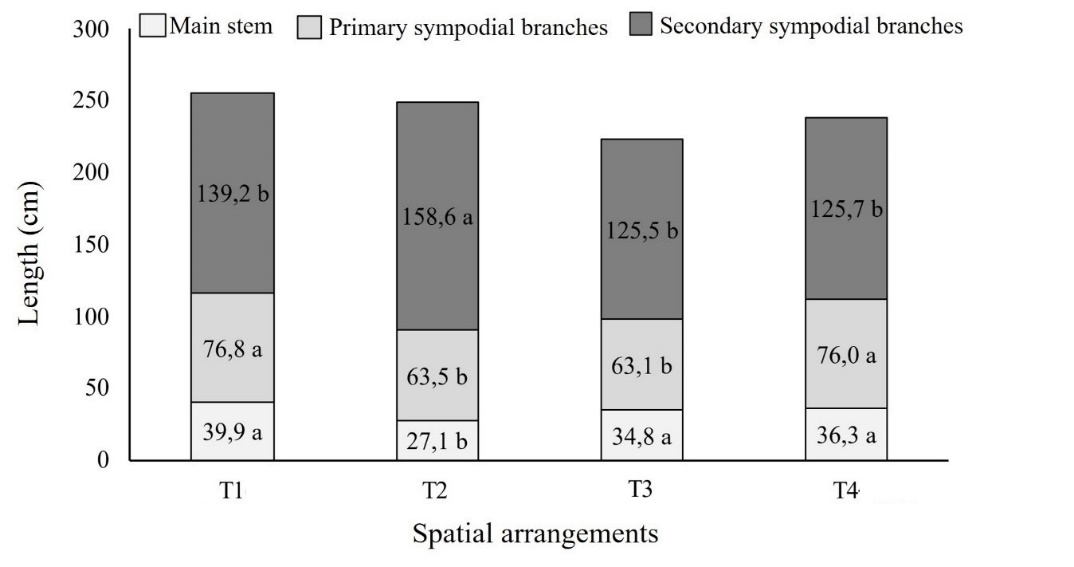
Fig 2. Lengths of the main stem, primary, and secondary sympodial branches of cassava (IAC 576-70) under the influence of cultivation systems. Pindorama, SP.
Table 4. Average root length and diameter, loss of non-commercial roots, and yield of cassava roots under the influence of different cultivation systems. Pindorama, SP.
| Cultivation systems | Root length (cm) | Root diameter (cm) | Commercial root losses (%) | Cassava Yield (Mg ha-¹) |
| T1 - Cassava monoculture | 29.20 b1 | 4.57 b | 21.94 bc | 41.66 a |
| T2 - Cassava + Peanuts between lines | 39.10 a | 4.15 c | 37.49 a | 33.50 b |
| T3 - Cassava + Peanuts in double lines | 39.30 a | 5.05 a | 13.65 c | 30.25 b |
| T4 - Cassava + Peanuts in dense double lines | 31.39 b | 4.71 b | 25.15 b | 28.92 b |
| F | 31.8331** | 34.8416** | 20.1015** | 44.7067** |
| MSD | 3.8851 | 0.2637 | 9.2646 | 7.03 |
| CV (%) | 5.95 | 3.04 | 20.08 | 12.44 |
1Averages followed by the same letter do not differ significantly by Tukey's test (p≤0.05).
systems did not show statistically different root diameter averages (p < 0.05), achieving, though, a root diameter 10.56% higher than the T2 treatment, which in turn presented a shorter root diameter (4.15 cm) (Table 4).
The increase in length and diameter of cassava roots indicates the number of carbohydrates translocated to the storage roots (Adeniji et al., 2020), and this simultaneously occurs with the biomass partitioning to the shoot (leaves, stem, and branches) throughout the plant's growth and development period. However, depending on the cultivar, the
photoassimilates metabolized by the plant in the first 90 to 180 DAP are mainly directed to the shoot, and, only after this period, which generally coincides with the dry matter maximum accumulation in the shoot, do the roots become the main drain of photoassimilates (El-Sharkawy, 2004; Souza et al., 2017).
Our results indicate that the cultivation systems did not affect the root growth phase but the storage one, which is partly supported by the estimated values of parameters tx, Am.tx/e, and DAPm from the Gompertz equation (Table 1) and growth rate curve (Fig 1). Higher values for the tx and
Table 5. Effect of different cultivation systems on the production and productivity of cassava and peanut crops. Pindorama, SP.
| Cultivation systems | Cassava | Peanut | ||
| Production (Mg. ha-¹) | Productivity (g. plant-1) |
Production (Mg. ha-¹) | Productivity (g. plant-1) |
|
| T1 - Cassava in monoculture | 41.66 a | 2500 b | - | - |
| T2 - Cassava + Peanuts between lines | 33.50 b | 1820 c | 2.220 b | 7.40 c |
| T3 - Cassava +Peanuts in double lines | 30.25 b | 4020 a | 1.875 b | 136.20 a |
| T4 - Cassava + Peanuts in dense double lines | 28.92 b | 2190 bc | 0.9585 c | 63.90 b |
| T5 - Peanuts in monoculture | - | - | 3.837 a1 | 129.10 a |
| F | 44.7067** | 44.7077** | 49.7490** | 42.1631** |
| MSD | 7.03 | 606.85 | 0.94 | 39.22 |
| CV (%) | 12.44 | 11.73 | 28.91 | 24.81 |
1 Averages followed by the same letter do not differ significantly by Tukey's test (p≤0.05).
Am.tx/e parameters indicate faster growth concerning plant height at the measurement time, especially after 90 DAP, when the plants showed satisfactory development. However, from the DAPm period onwards (Table 1), the cassava plants grown in the T2 and T4 systems showed a sharper drop in growth rate (tx and Am.tx/e) compared to the T1 and T3 systems (Fig 1), suggesting the cassava plants were more intensely affected by competing growth factors, which impacted tuber bulking and consequently plant productivity.
Commercial root losses were the highest in the treatment with peanuts planted directly between the lines of the cassava (T2), followed by the treatment with cassava planted in dense double rows (T4), where the proximity of the cassava plants enhanced interspecific competition, or at least a partial one, with the intercropped peanut. However, the commercial root losses in the treatment T4 were not significantly different from T1 (p < 0.05), which in turn did not differ statistically from T3 (p < 0.05) either and were probably due to higher competition for physical space, water, and nutrients, especially during tuber bulking. Moreover, the trichotomous branching pattern of the cassava cultivar (IAC 576-70) may have influenced the partitioning of resources in the plant. According to Fagundes et al. (2009), cassava plants with trichotomous branching have a higher branch number on the upper secondary sympodial branches. This pattern occurs because each branching emits three new stems, requiring significant resources for these structures to grow and develop (Fagundes et al., 2009).
Although the results of this study did not detect differences in the number of branches (Table 2), the cassava cultivar IAC 576-70 was affected concerning the length of the secondary sympodial ones (Fig 2). This length increase indicates a higher resource demand to grow these structures, resulting in lower plant productivity, especially in denser treatment (T2), which had longer secondary sympodial branches (Fig 2). According to Souza et al. (2017), photoassimilates can be directed to the cassava shoot to support the development of new structures, such as leaves and branches, during the period the plant most accumulates dry matter, which may, therefore, have caused the longer branch growth observed in T2.
Concerning cassava productivity, the monoculture system (T1) provided the highest productivity (41.66 Mg ha-1), 25.85% higher than the cultivation systems T2, T3 and T4. However, these treatments (T2 and T3) did not differ from each other (p < 0.05), with an average variation of ~31 Mg ha-1 (Table 4). These results partially corroborate previous studies, such as those by Dapaah et al. (2003), Albuquerque et al. (2012), Akpan & Ikeh (2018), Horacio et al. (2019), and Mansaray et al. (2022). Legodi & Ogola (2020) found that cassava root production in monoculture was also higher when compared to cassava production intercropped with cowpeas, chickpeas, and Bambara groundnuts (Vigna subterranea). Moreover, the authors observed that cowpeas were more competitive than chickpeas and Bambara
groundnuts. They point out that the cowpea plant size and its ability to acquire higher aerial biomass are crucial factors, negatively influencing the development of cassava and resulting in lower root production.
As for the productivity and production per plant of the crops, all the cultivation systems negatively affected the productivity of the two crops compared to their monoculture (Table 5). Cassava showed a significant decrease in productivity only when monoculture and mixed crops were considered, with the spatial intercropping arrangements not differing. For the peanut arrangement between the rows (T2), direct competition for the production factors may have been the cause of the decrease in productivity, compared to the cassava monoculture (T1), which respected the same spacing.
Spacing between plant rows is a significant factor to consider in plantations where the spacing between plants is small and affects productivity. When looking at individual yields, the plants that went through less intraspecific competition produced a higher root mass individually, as occurred in the treatment with alternating double rows of cassava and peanuts (T3), in which the cassava plants could overlap to receive light in the peanut rows with more physical space for the roots to grow as well, especially after the peanut harvest, in which case the average yield per plant was higher than in the monoculture (T1) (Table 5).
In the treatments with less dense rows, competition for production factors did not affect crop production per plant compared to monoculture (T3). However, competition for resources, such as water, seriously affected individual plant production, especially in treatments T2 and T4. These results are comparable to reports by Albuquerque et al. (2012) and Legodi and Ogola (2020), who found a reduction in the production of common beans, cowpeas, chickpeas, and Bambara peanuts, when intercropped with cassava, compared to monocropping. The authors point out that the cultivation arrangement and the species intercropped can alter the behavior of the crops due to competition for resources.
As for partial LUE, the intercropping between cassava and peanuts in the spatial arrangements applied showed lower indices when compared to the values of their respective monocultures (Table 6). However, when comparing the partial LUE index values of T2, T3, and T4 cultivation systems for cassava, the partial LUE values differed by 20%, 27%, and 31%, respectively, from the cassava monoculture. However, regarding the peanut crop, the difference increased to 42%, 51%, and 75% for the T2, T3, and T4 cropping systems, respectively, compared to the peanut monoculture (Table 6). Partial LUE indices with values close to those observed in monocropping systems indicate higher competitiveness of the cassava crop concerning the peanut. Rós and São João (2016) observed similar results for the cassava crop when they evaluated the partial LUE in the cultivation system involving cassava intercropped with sweet potato.
Table 6. Land use efficiency (LUE) of the cassava-peanut intercropping under the influence of different cultivation systems.
| Cultivation systems | Partial LUE | Total LUE | |
| Cassava | Peanuts | ||
| T1 - Cassava in monoculture | 1 | - | 1 |
| T2 - Cassava + Peanuts between lines | 0.80 | 0.58 | 1.38 |
| T3 - Cassava +Peanuts in double lines | 0.73 | 0.49 | 1.21 |
| T4 - Cassava + Peanuts in dense double lines | 0.69 | 0.25 | 0.94 |
| T5 - Peanuts in monoculture | - | 1 | 1 |
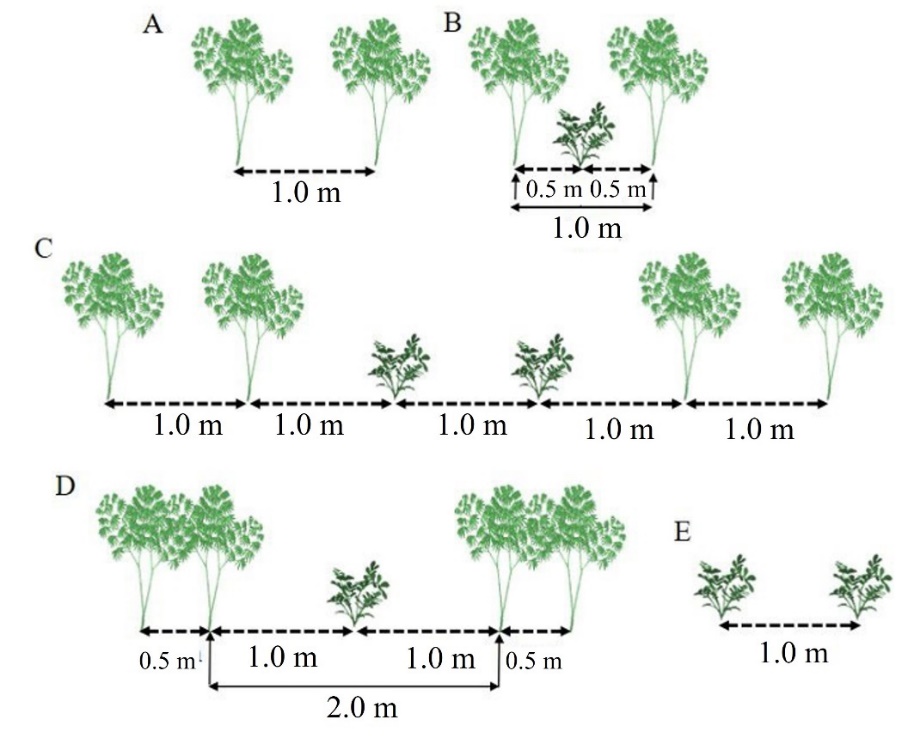
Fig 3. Schematic representation of cassava and peanut cultivation patterns. A: Cassava monoculture, with rows spaced 1 m apart and 0.6 m between plants in the row (T1); B: Cassava+peanut intercropping, with peanut rows spaced 0.50 m from the cassava rows (T2); C: Cassava+peanut intercropping, with double rows of cassava and double rows of peanuts, spaced 1.0 m apart (T3); D: Cassava+peanut intercropping, with a double row of cassava spaced 2.0 m apart and a single row of peanuts spaced 1.0 cm from the double row of cassava (T4); E: peanut monoculture, with rows spaced 1.0 m apart (T5). Pindorama, SP.
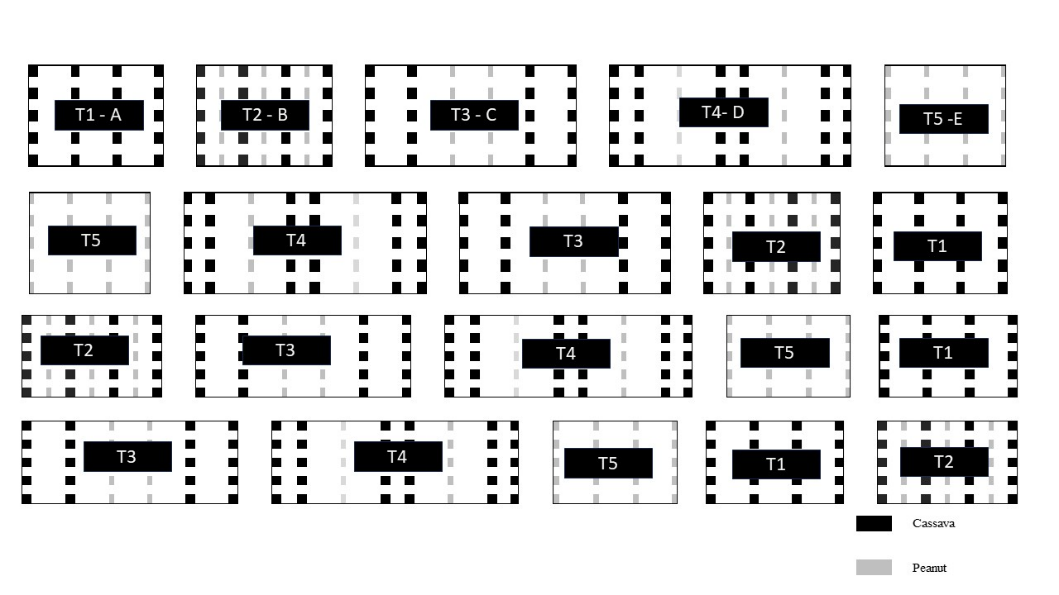
Fig 4. Distribution of treatments in the experimental plots. T1: Cassava monoculture, with rows spaced 1 m apart and 0.6 m between plants in the row; T2: Cassava+peanut intercropping, with peanut rows spaced 0.50 m from the cassava rows; T3: Cassava+peanut intercropping, with double rows of cassava and double rows of peanuts, spaced 1.0 m apart; T4: Cassava+peanut intercropping, with a double row of cassava spaced 2.0 m apart and a single row of peanuts spaced 1.0 cm from the double row of cassava; T5: peanut monoculture, with rows spaced 1.0 m apart. Pindorama, SP.
Regarding total LUE, the intercropping of cassava and peanuts, with peanuts planted between the cassava rows (T2), had a 38%-greater total LUE than monocropping. For intercropped double rows (T3), the gain in LUE was 21% (Table 6). This shows that even if the productivity per unit area of intercropped cultures was lower (Tables 4 and 5) than the monoculture systems, the LUE with intercropping is
higher than with isolated monocultures, provided the right arrangement is chosen. This is supported by the results of the partial LUE index for the T2 and T3 cropping systems that displayed less difference (20%, 27%, 42%, and 51%) than their monocultures. Moreover, the results show that the two species, when grown in monoculture, require between 21 and 38% more land to produce the same amount of grain and roots compared to the intercropped area. Similar results have been reported for cassava and common bean intercrops (Albuquerque et al., 2012); cassava and sweet potato (Rós and São João, 2016); cassava and cowpea, chickpea, and Bambara
peanut intercrops (Legodi and Ogola, 2020) and between cassava and cowpea intercrops (Amoako et al., 2022).
According to Rós and São João (2016), intercropping and combining different spatial arrangements of species can affect the productivity of the intercropped cultures, leading to an increase in productivity per unit area. These authors also add that it is essential to relate the productivity of each crop to its potential commercial value at harvest time so that the feasibility of using the land efficiently can increase income.
Thus, even though the intercropping system affects crop yields, the overall yearly cultivation can bring the farmer crop productivity if the selection of the arrangement is appropriate. In addition, the beneficial effects of intercropping with legumes can improve soil fertility for subsequent crops, reduce the incidence of weeds (Albuquerque et al., 2012; Mudare et al., 2022), and benefit small producers who wish to maximize productivity in limited areas, as well as reducing cultivation costs (Alves et al., 2010).
Material and Methods
Experiment location and plant materials
The experiment was conducted at the Agência Paulista de Tecnologia do Agronegócio (APTA) - North Central Regional Hub, in Pindorama, São Paulo, Brazil. The area with Eutrophic Red-Yellow Argisol soil, suitable for cassava and peanuts, has historically been planted with cassava. The IAC 576-70 cultivar was chosen due to its predominance and prominence in the cultivation of sweet cassava in São Paulo, representing 90% of the area in the region. Recognized for its high productivity and culinary qualities, this cultivar is adapted to local agricultural practices (Silva et al., 2021). The peanut cultivar adopted was the IAC Caiapó, notable for its resistance to leaf diseases such as black spots and rust (Godoy et al., 1999).
Experimental design
The experimental design was a randomized block with 05 treatments and 05 replications, totaling 25 plots. Cassava (IAC 576-70) and peanut (IAC Caiapó) were planted asynchronously, with the peanuts being sown 40 days after the cassava plantation to ensure the latter establishment. The treatments (T) included cassava in monoculture (T1) with six rows, the central four of which were considered the plot. In T2, the peanut line was sown between the cassava lines. The T3 treatment had double rows of both cassava and peanuts alternated. T4 had cassava rows in double dense planting, with a single row of peanuts between them. The peanuts in the monoculture treatment (T5) had six rows. Figure 3 schematically outlines the spacing of the cultivation systems and Figure 4 describes the distribution of treatments in the experimental plots.
Cassava vegetative growth
From the 60th day of planting the cassava and the 10th day of coexistence between the species, the height of the cassava plants was measured monthly until the cassava was harvested 270 DAP, when 10 plants were finally selected for complementary measurements concerning the heights of their primary branching (TB1) and secondary sympodial branch (SB2), along with the lengths of the main stem and primary and secondary sympodial branch, using a millimeter ruler, with the % branching obtained from this data.
Crop production, cassava root quality, and land use efficiency index
At 150 DAP, the peanuts crop at the end of its cycle was harvested and measured with 13% moisture for its yield. At 270 DAP, it was the cassava crop turn, and the entire experimental plot was assessed for yielding, separating the commercial quality roots from the ones considered losses. The cassava and peanut yield data allowed the LUE index to be calculated.
Statistical analysis
The growth curves of the cassava plants obtained every two weeks were estimated using the Gompertz equation (1825), as follows:
\(H = Hm.e^{{- e}^{- mr(t - DAPm)}}\)
Where: H = plant height (m) at time t for each cassava spatial arrangement, in monoculture or intercropped; Hm = maximum potential height; mr = maximum relative growth rate at the inflection point of the curve (m.m-1 per day); DAPm = time (days after planting) at which the growth rate is maximum. Based on the estimated equations, the growth rates (g/day) were calculated as a function of time (t), using the derivative of the Gompertz equations. The characteristics of the Gompertz curve revolve around the inflection point at which the growth rate is maximum. So, in this case, the time at which the inflection point occurs is given by the DAPm parameter of the function. At this point, height equals A/e (36.8% of the maximum), and the growth rate equals A.B/e.
The vegetative data collected at harvest time (plant height, height of sympodial branches, lengths of the main stem and primary and secondary sympodial branches, diameter of the main stem, and % branching), productive data (yield and production per cassava and groundnut plant) and the qualitative data (root length, root diameter and losses) were subjected to the Shapiro-Wilk normality test, qualitative (length, diameter and losses of roots), were subjected to the Shapiro-Wilk normality test, with no need for transformation. The data was then subjected to ANOVA using the F test (p≤0.05), and the means were compared using the Tukey test (p≤0.05).
Peanut and cassava production data were used to determine land use efficiency. The LUE index was defined as the relative area of land under single planting conditions required to provide the yields achieved in intercropping and is calculated by the following expression: LUE = Ic + Ip, where Ic and Ip represent the efficient land use indices of the crops. The formulas Ic = Ic. (Mc)-¹ and Ip = Ip. (Mp)-¹, where Ic and Ip are the cassava and peanut yields in intercropping, and Mm and Ma are their yields in monocropping (Mead and Willey, 1980).
Conclusions
Cultivation systems involving intercropping and the spatial arrangement combinations between cassava and peanuts affect the vegetative growth of cassava plants regarding height, branching, length, root diameter, and plant productivity, especially when using spatial arrangements that increase interspecific and intraspecific competition between plants.
Planting cassava and peanuts in less dense arrangements offers more advantageous conditions for the cassava root growth concerning length and diameter, resulting in lower root losses.
Intercropping cassava with peanuts in denser arrangements is advantageous for land use efficiency (LUE).
Acknowledgments
The authors would like to thank the Agência Paulista de Tecnologia dos Agronegócios (APTA, Pindorama-SP) for supporting the development of this research, the Postgraduate Program in Agronomy (POSAGRO) at the Federal University of Roraima and the National Council for Scientific and Technological Development (CNPq).
Authors Contributions
Maria Beatriz Bernardes Soares: Experiment design, guidance, data collection, initial scientific writing, statistical analysis and translation into English; Everton Luis Finoto and José de Anchieta Alves de Albuquerque: Guidance and review of scientific writing; Glauber Ferreira Barreto, João Luiz Lopes Monteiro Neto, José Maria Arcanjo Alves, Richard Alcides Molina Alvarez, Laura Soliane Cruz Braz, Anderson Carlos de Melo Gonçalves and Edgley Soares da Silva: review of the scientific writing.
References
Adeniji OT, Okonji CJ, Adewale DB, Badmus AA, Adewoyin OB, Baiyeri SO, Iseghohi IO, Sabo E, Eromosele G, Odetoye F (2020) Evaluation of Agronomic, Root Metric and Participatory Assessment of Root Quality Attributes in Orange- and White-Fleshed Cassava Clones. J Food Agric Environ. 7:1-16.
Agegnehu G, Ghizaw A, Sinebo W (2006) Yield performance and land-use efficiency of barley and faba bean mixed cropping in Ethiopian highlands. Eur J Agron. 25:202-207.
Akpan EA, Ikeh AO (2018) Growth and yield response of cassava (Manihot esculenta Crantz) varieties to different spacing in Uyo, Southeastern Nigeria. J. Agric. Crop Res. 6:19-27.
Albuquerque JAA, Oliva LSC, Alves JMA, Uchôa SCP, Melo DA (2015) Cultivation of cassava and cowpea in intercropping systems held in Roraima’s Savannah, Brazil. Rev Cienc Agron. 46:388-395.
Albuquerque JAA, Sediyama T, Alves JMA, Silva AA, Uchôa SCP (2012) Cultivo de mandioca e feijão em sistemas consorciados realizado em Coimbra, Minas Gerais, Brasil. Rev Cienc Agron, 43:532-538.
Alves JMA, Araújo NP, Uchôa SCP, Albuquerque JDAA, Silva AJ, Rodrigues GS, Silva DCO (2010) Avaliação agroeconômica da produção de cultivares de feijão-caupi em consórcio com cultivares de mandioca em Roraima. Rev Agro@mbiente On-Line, 3:15.
Amoako OA, Adjebeng-Danquah J, Agyare RY, Tengey TK, Puozaa DK, Kassim BY (2022) Growth and yield performance of cassava under different plant population densities of two cowpea varieties in a cassava/cowpea intercrop. Int J Agric Technol. 18:1917-1936.
Botelho F, Peixoto CP, Almeida AT, Botelho GLP (2020) Methods of cultivation and technical coefficients of peanuts in the Bahian Recôncavo. Arquivos Do Instituto Biológico, 87:1-9.
Dapaah HK, Asafu-agyei SA, Ennin SA, Yamoah C (2003) Yield stability of cassava, maize, soya bean and cowpea intercrops. J Agr Sci. 140:3-82.
El-Sharkawy MA (2004) Cassava biology and physiology. Plant Molecular Biology, 56: 481–501.
Fagundes LK, Streck NA, Lopes SJ, Rosa HT, Walter LW, Zanon AJ (2009) Desenvolvimento vegetativo em diferentes hastes da planta de mandioca em função da época de plantio. Cienc Rural. 39:657–663.
Gabriel LF, Streck NA, Roberti DR, Chielle ZG, Uhlmann LO, Silva MR, Silva SD (2014) Simulating Cassava Growth and Yield under Potential Conditions in Southern Brazil. Crop Ecology & Physiology, 106:1119-1137.
Godoy IJ, Moraes SA, Siqueira WJ, Pereira JCVNA, Martins ALM, Paulo EM (1999) Produtividade, estabilidade e adaptabilidade de cultivares de amendoim em três níveis de controle de doenças foliares. Pesqui Agropecu Bras. 34:1183–1191.
Horacio EH, Mota JH, Teixeira IR (2019) Monocultivo e cultivo consorciado de feijão e mandioca. Sci Agrar. 18(2):132-138.
IBGE - Instituto Brasileiro de Geografia e Estatística (2022) Levantamento Sistemático da Produção Agrícola. Rio de Janeiro-RJ, Brasil.
Legodi KD, Ogola JBO (2020) Cassava-legume intercrop: I. Effects of relative planting dates of legumes on cassava productivity. Plant Sci. 70:150-157.
Leihner D (ed) (1983) Management and evaluation of intercropping systems with cassava. Centrol International de Agricultural Tropical, Cali, Colombia.
Lima CA, Montenegro AAA, Santos TEM, Andrade EM, Monteiro ALN (2015) Agricultural practices in the cultivation of cassava and the relation to runoff, and soil and water loss. Rev Cienc Agron. 46:697-706.
Mahakosee S, Jogloy S, Vorasoot N, Theerakulpisut P, Toomsan B, Holbrook CC, Kvien CK, Banterng P (2022) Light Interception and Radiation Use Efficiency of Three Cassava Genotypes with Different Plant Types and Seasonal Variations. Agron. 12:2888.
Maitra S, Hossain A, Brestic M, Skalicky M, Ondrisik P, Gitari H, Brahmachari K, Shankar T, Bhadra P, Palai JB, Jena J, Bhattacharya U, Duvvada SK, Lalichetti S, Sairam M (2021) Intercropping - a low input agricultural strategy for food and environmental security. Agron. 11:343.
Mansaray A, Karim AB, Yormah TBR, Conteh AR, Yila KM (2022) Effect of Time of Introduction of Legumes into Cassava on the Productivity of Cassava in Cassava-Legume based Intercropping Systems. Asian J Adv Agric Res, 18:1-15.
Mead R, Willey RW (1980) The concept of a land equivalent ratio and advantages in yields for intercropping. Exp Agr. 6:217-228.
Mudare S, Kanomanyanga J, Jiao X, Mabasa S, Lamichhane JR, Jing J, Cong W (2022) Yield and fertilizer benefits of maize/grain legume intercropping in China and Africa: A meta-analysis. Agron Sustain Dev. 42:1-17.
Oliveira EC, Almeida LHC, Zucareli C, Valle TL, Souza JRP, Miglioranza É (2019) Análise do crescimento de mandioca em diferentes épocas de colheita e densidades de plantio. Semin Cienc Agrar. 40:113–126.
Peoples MB, Giller KE, Jensen ES, Herridge DF (2021) Quantifying country-to-global scale nitrogen fixation for grain legumes: I. Reliance on nitrogen fixation of soybean, groundnut and pulses. Plant Soil, 469:1–14.
Reichert JM, Fontanela E, Awe GO, Fasinmirin JT (2021) Is cassava yield affected by inverting tillage, chiseling, or additional compaction of no-till sandy-loam soil? Rev Bras Cienc Solo. 45.
Rós AB, João RES (2016) Agronomic performance and land equivalent ration in cassava and sweet potato arrangements plants. Rev. Ceres, 63:517-522.
Rubiano-Rodríguez JA, Cordero-Cordero CC (2019) Épocas críticas de competencia de arvenses en cultivo de yuca en el Caribe seco colombiano. Temas Agrarios, 24:108-118.
Silva DCO, Alves JMA, Uchôa SCP, Sousa AA, Barreto GFS, Silva CN (2017) Curvas de crescimento de plantas de mandioca submetidas a doses de potássio. Rev Cienc Agr, 60:158-165.
Silva MA, Zanetti S, Santos LH, Pereira PLF (2021) Nutritional status, yield components, and yield of cassava as influenced by phenological stages and water regimes. J Plant Nutr. 44:1–16.
Souza AP, Massenburg LN, Jaiswal D, Cheng S, Shekar R, Long SP (2017) Rooting for cassava: insights into photosynthesis and associated physiology as a route to improve yield potential. New Phytol. 213:50-65.
Streck NA, Pinheiro DG, Zanon AJ, Gabriel LF, Rocha TSM, Souza AT, Silva MR (2014) Efeito do espaçamento de plantio no crescimento, desenvolvimento e produtividade da mandioca em ambiente subtropical. Bragantia. 73:407–415.