Aust J Crop Sci. 19(01):18-24 (2025) | ISSN:1835-2707
https://doi.org/10.21475/ajcs.25.19.01.p135
Novel genetic resources: Rice varieties for improving direct seeding genotypes
Anuson Thanasi1, Samart Wanchana2 and Sureeporn Kate-ngam*1
1Department of Agronomy, Faculty of Agriculture, Ubon Ratchathani University, Warin Chamrap, Ubon Ratchathani 34190 Thailand
2National Center for Genetic Engineering and Biotechnology (BIOTEC), 113 Thailand Science Park, Phahonyothin Road, Khlong Nueng, Khlong Luang, Pathum Thani 12120 Thailand
*Corresponding author: Sureeporn Kate-ngam 
Abstract: Early seedling vigor is an essential characteristic for direct seeded rice. This study aimed to evaluate thirty-one rice varieties, categorized into three ecotypes (lowland, upland, and deep-water cultivars), to identify traits associated with early seedling vigor under direct seeding conditions. Each evaluation was conducted using three biological replicates. The experiments comprised of two parts: (a) deep-sowing tolerance and anaerobic germination tests and (b) aerobic germination conditions as a control. Early seedling vigor traits in rice, including mesocotyl length (MS), shoot length (SL), coleoptile length from deep-sowing experiment (COD), shoot weight (SW), coleoptile length from the anaerobic germination experiment (COA), coleoptile length under normal conditions (CON), the anaerobic response index (AI), and seedling vigor (SV), were measured. The results demonstrated significant heritability across all traits. Principal component analysis identified CON and COD as the key variables influencing early seedling vigor traits. By using a multi-traits selection index, that considered eight important traits, U-19 (Plawng Aew), U-11 (Chum Phae 60), U-31 (Hawm Bao Rai), U-5 (Pin Gaew 56), and U-2 (Prachinburi 2) varieties were prioritized as important genetic resources for the enhancement of direct-seeding rice varieties. This research provides valuable insights into the traits associated with early seedling vigor. It is crucial for improving rice varieties that are widely adaptable to direct seeding conditions. The identified varieties showed promising for advancing breeding projects aimed at increasing the efficiency and sustainability of direct-seeding rice production systems.
Keywords: Rice, Direct seeding, Seedling vigor, Deep-sowing tolerance, Anaerobic tolerance
Introduction
Rice is a vital crop for human food security with a predominant method of production involving transplanting. However, challenges such as climate change-induced water scarcity, rising labor costs, and expanding urbanization have rendered transplanting unsustainable (Edzesi et al., 2023). Recently, there has been a noticeable shift from transplanting to direct seeding in rice production (Castano et al., 2021).
Direct seeding, particularly prevalent in Southeast Asia (Hsu and Tung, 2015; Yang et al., 2022), addresses some of the limitations associated with transplanting. However, it introduces new challenges, such as poor emergence, low early seedling vigor, and suboptimal establishment (Angaji et al., 2010; Ghosal et al., 2019; Wang et al., 2021). Various factors, including environmental conditions and genetics, contribute to these issues. For instance, flooding during the germination stage caused by unpredictable rainfall and variations in seeding depth due to tractor machinery can hinder the growth and development of rice (Hsu and Tung, 2015; Zhao et al., 2018). Therefore, there is a pressing need to develop specific rice varieties tailored for direct seeding to overcome these challenges (Zhan et al., 2020).
Ensuring the early vigor of seedlings, which is refereed to deep sowing tolerance and anaerobic germination tolerance, is a critical agronomic attribute for developing rice varieties suited to direct seeding. Successful direct seeding requires uniform and rapid germination, along with robust establishment, even under diverse environmental stresses (Mahender et al., 2015). Several studies have identified mesocotyl and coleoptile elongation as key traits for rice adaptation to unpredictable rainfall and varying seeding depths, conditions that often lead to poor early seedling vigor (Edzesi et al., 2023; Hsu and Tung, 2015; Lee et al., 2017; Nghi et al., 2019; Xiong et al., 2017; Zhan et al., 2020). The elongation of the mesocotyl and coleoptile is influenced by a range of genes distributed across the rice genome, as identified through quantitative trait loci (QTL) mapping and genome-wide association studies (GWAS) (Edzesi et al., 2023; Jang et al., 2021; Lee et al., 2012; Lu et al., 2016; Wang et al., 2021; Wu et al., 2015; Zhang et al., 2023). Furthermore, phytohormones play a significant role in this process, particularly the interaction between ethylene and jasmonic acid, which has been shown to contribute to elongation (Xiong et al., 2017). Other factors, such as strigolactones (Hu et al., 2010) and auxin (Nghi et al., 2021), also exert a considerable influence. These findings collectively deepen our understanding of the complex regulatory networks governing mesocotyl and coleoptile elongation in rice, thereby aiding in the development of varieties that are better suited to direct seeding practices.
The objective of this study was to screen thirty-one varieties, comprising of upland, lowland, and deep-water cultivars, to identify traits associated with early seedling vigor. These traits are essential for identifying new genetic resources that can be utilized in breeding programs aimed at enhancing rice varieties for direct seeding.
Results and Discussion
Evaluation of deep sowing tolerance and anaerobic germination tolerance
In this study, anaerobic germination and the deep-sowing tolerance of 31 rice varieties were evaluated by collecting data at 7 and 10 days after treatment, respectively. All agronomic traits,
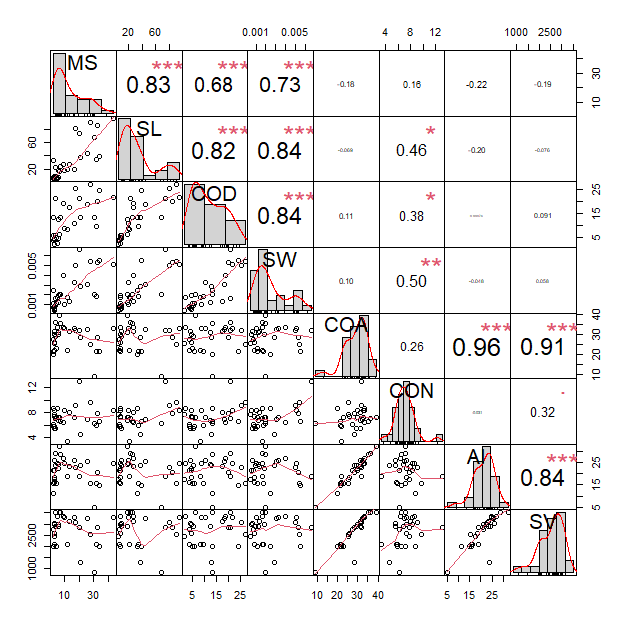
Fig 1. The correlation analysis presented here delves into the traits pivotal for early seedling vigor in rice. There were eight examined parameters include mesocotyl length (MS), shoot length (SL), coleoptile length of deep-sowing experiment (COD), shoot weight (SW), coleoptile length of anaerobic germination experiment (COA), coleoptile length of normal condition (CON), anaerobic response index (AI), and seedling vigor (SV). The statistical significance of these correlations is marked by * for p < 0.05 and *** for p < 0.001. Twelve significant positive correlations were identified. Remarkably, MS exhibited a strong positive correlation with SL, COD, and SW, while SV notably displayed correlation with AI and COA.
including mesocotyl length (MS), shoot length (SL), coleoptile length from the deep-sowing experiment (COD), shoot weight (SW), coleoptile length from the anaerobic germination experiment (COA), coleoptile length under normal conditions (CON), anaerobic response index (AI), and seedling vigor (SV), were found to be highly significant (p < 0.000) (Table 1). The MS ranged from 2.13 mm to 43.95 mm, with an average value of 13.30 mm; the SL ranged from 5.04 mm to 95.31 mm, with an average value of 30.94 mm; the COD ranged from 1.75 mm to 27.08 mm, with an average value of 11.95 mm; and the SW ranged from 0.0004 g to 0.0066 g, with an average value of 0.0024 g. Meanwhile, the COA ranged from 22.05 mm to 31.75 mm, with an average value of 28.25 mm; the CON ranged from 3.34 mm to 13.05 mm, with an average value of 7.19 mm; the AI ranged from 4.99 to 32.18, with an average value of 21.13; and the SV ranged from 825.06 to 3530.65, with an average value of 2701.39. The broad-sense heritability varied from 79% for AI to 98% for SL, with COA and MS showing heritability values of 80% and 97%, respectively (Table 1). These findings are consistent with previous reports (Uzair et al., 2022). Heritability was categorized as low (below 30%), medium (30% - 60%), or high (above 60%). In this study, all parameters exhibited high heritability except for SW, suggesting that selection for these traits would be effective due to minimal environmental influence. Furthermore, these results may prove useful in identifying the genetic factors that are predictive of breeding program success (Hossain et al., 2021). The coefficient of variation ranged from 10.16% for COA to 29.63% for SW (Table 2).
Correlation analysis and principal component analysis (PCA)
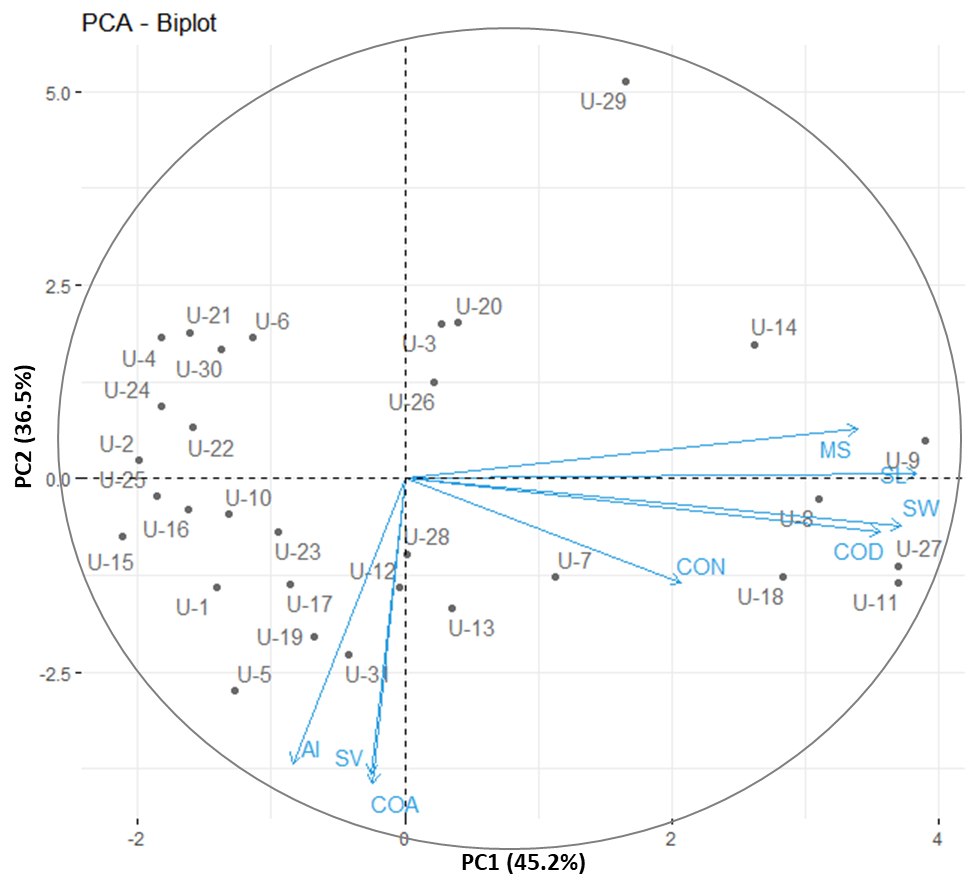
The correlation analysis indicated that mesocotyl length (MS) exhibited a significantly positive correlation with shoot length (SL) (r = 0.83***), coleoptile length from the deep-sowing experiment (COD) (r = 0.82***), and shoot weight (SW) (r =
0.73***). Additionally, the coleoptile length from the anaerobic germination experiment (COA) showed a significantly positive correlation with the anaerobic response index (AI) (r = 0.96***) and seedling vigor (SV) (r = 0.91***) (Fig. 1). The first two principal components for early agronomic traits had eigenvalues greater than one, collectively accounting for 81.70% of the total variance. Principal Component 1 (PC1) explained 45.20% of the variance, while Principal Component 2 (PC2) explained 36.50% of the cumulative variance in early agronomic traits of rice (Fig. 2). PC1 was positively correlated with increases in SL, MS, SW, COD and CON, while PC2 was highly negative correlated with COA, SV and AI. The upper right quadrant of Fig. 2 shows that U-9 was the most superior cultivar, whereas cultivars with lower growth at the early stage, such as U-15 and U-2, were located in the left quadrant. In this study, Principal component analysis (PCA) was employed to discern patterns and identify key variables shaping our dataset. Notably, the variable denoted as coleoptile length under normal conditions (CON) emerged as a consistently influential factor, displaying substantial loadings across multiple principal components (PC1, PC2, PC3, PC4, PC5, PC7, and PC8). Furthermore, COD also demonstrated significant loadings across several components (PC1, PC3, PC4, PC6, and PC7) (Table 3), underscoring its role as another influential factor with broad-reaching effects. Prioritizing traits linked to CON and COD could confer strategic advantages in optimal trait selection, given their extensive impact across various aspects of the dataset. This data-driven approach through PCA provides valuable insights that facilitate informed decision-making in the prioritization of traits (Ashfaq et al., 2023; Uzair et al., 2022).
The elongation of the coleoptile and mesocotyl is crucial for enhancing seedling establishment during direct seeding. Under submergence stress during the germination stage, rapid coleoptile elongation is a key trait to ensure productive rice growth (Barik et al., 2019; Castano et al., 2021; Ghosal et al., 2019; Ismail et al., 2012; Shin et al., 2020; Su et al., 2021; Toledo et al.,
Table 1. Analysis of variance of related traits associated with early seedling vigor refer to deep sowing tolerant and anaerobic germination tolerant parameters in 31 rice varieties.
| Traits | df | MS | EMS | Min | Max | Mean | H2 (%) |
|---|---|---|---|---|---|---|---|
| MS | 30 | 427.50** | 5.58 | 2.13 | 43.95 | 13.30 | 97 |
| SL | 30 | 2146.12** | 13.62 | 5.04 | 95.31 | 30.94 | 98 |
| COD | 30 | 174.54** | 6.16 | 1.75 | 27.08 | 11.95 | 93 |
| SW | 30 | 0.00** | 0.00 | 0.0004 | 0.0066 | 0.0024 | NA |
| COA | 30 | 79.84** | 8.73 | 22.05 | 31.75 | 28.25 | 80 |
| CON | 30 | 8.10** | 0.76 | 3.34 | 13.05 | 7.19 | 82 |
| AI | 30 | 76.35** | 8.86 | 4.99 | 32.18 | 21.13 | 79 |
| SV | 30 | 1033040.96** | 103994.77 | 825.06 | 3530.65 | 2701.39 | 81 |
Note: MS = mesocotyl length (mm), SL = shoot length (mm), COD = Coleoptile length of deep-sowing experiment (mm), SW = shoot weight (g), COA = Coleoptile length of anaerobic germination experiment (mm), CON = Coleoptile length of normal condition (mm), AI = anaerobic response index, SV = seedling vigor, df = degree of freedom, MS = mean of sum square, EMS = error of mean square, Min = minimum, Max = maximum, H2 = broad-sense heritability, ** indicates the significance levels at p < 0.01.
Fig 2. The Principal Component Analysis (PCA) biplot, based on early seedling vigor parameters and the projection of variables, revealed that PC1 and PC2 together captured 81.7% of the variation, primarily affecting seedling length. These principal components (PCs) serve as indicators for seedling length (SL) and seedling weight (SW). PC1 showed that seedling length could be a useful trait for selecting cultivars. (MS = mesocotyl length, SL = shoot length, COD = coleoptile length of deep-sowing experiment, SW = shoot weight, COA = coleoptile length of anaerobic germination experiment, CON = coleoptile length of normal condition, AI = anaerobic response index, SV = seedling vigor).
2015). Similarly, mesocotyl elongation is an important trait for deep-sowing tolerance, as it accelerates the establishment and emergence rate of rice seedlings under stress conditions induced by machinery, such as tractors (Lee et al., 2017; Simon et al., 2012; Wu et al., 2015; Zhao et al., 2018).
Hierarchical clustering dendrogram
The hierarchical clustering dendrogram revealed that the thirty-one varieties were grouped into three distinct clusters. Cluster I exhibited a complex structure, characterized by greater coleoptile elongation under submergence stress during the germination stage, which is consistent with the PCA results for PC2. Cluster II reveals that the majority of the varieties in this cluster belong to the lowland rice ecotype. Cluster III comprised of only two varieties, both identified as upland rice cultivars: U-26 and U-29 (Fig. 3).
Selection index
Eight traits identified as important for enhancing rice genotypes for direct seeding were utilized in the selection index. Many researchers have reported that focusing on a single trait may
ultimately lead to the neglect of other crucial traits. Multi-traits selection enhances the effectiveness of germplasm selection and has been successful in various breeding projects (Anshori et al., 2019; Candido et al., 2020; Jahufer and Casler, 2015; Mar et al., 2020; Ramos et al., 2021; Smiderle et al., 2019; Wang et al., 2019). The rankings of the 31 varieties, according to the Smith-Hazel index with a selection pressure of 20%, are illustrated in Fig 4.
The selection index was developed by combining traits associated with deep sowing, such as MS, SL, and SW. Using this index, five varieties U-11 (Chum Phae 60), U-9 (Khi Tom Gamnan), U-8 (Niew san pah tawng), U-18 (Khao Pahn Luang), and U-29 (Beu PaTaw) were selected (Fig. 4a). Among these, U-11 is an improved rice variety, which was resulted from a cross between Gam Pai 41 (male parent) and Leuang Tawng 78 (female parent). The female parent, Leuang Tawng 78, is an upland rice variety traditionally cultivated in Northern Thailand, while U-29 is also classified as an upland rice variety. This indicates that traits conferring deep sowing tolerance are predominantly found in upland rice germplasm. These findings align with those of Jang et al. (2021), who reported that upland rice varieties generally develop longer mesocotyls than lowland rice varieties, a trait
Table 2. Related traits associated with early seedling vigor parameters in 31 rice varieties.
| Entries | Varieties | MS | SL | COD | SW | COA | CON | AI | SV |
|---|---|---|---|---|---|---|---|---|---|
| U-1 | RD45 | 5.19g | 7.00ef | 2.25h | 0.0017b-f | 33.29abc | 8.62abc | 24.67abc | 3329.16a |
| U-2 | Prachinburi 2 | 2.13g | 5.37ef | 3.75fgh | 0.0007f | 28.63abc | 6.07bcd | 22.56abc | 2862.53ab |
| U-3 | RD19 | 2.70g | 39.23a-f | 13.38a-h | 0.0025 b-f | 18.77cd | 8.75abc | 10.02cd | 1876.63abc |
| U-4 | Plai Ngahm Prachinburi | 2.97g | 4.04f | 3.03gh | 0.0008f | 20.06bcd | 6.20bcd | 13.86bcd | 2006.25abc |
| U-5 | Pin Gaew 56 | 6.55fg | 17.24c-f | 6.12d-h | 0.0020 b-f | 39.38a | 7.20bcd | 32.18a | 3530.65a |
| U-6 | Gaen Jan | 12.08d-g | 20.41c-f | 8.33c-h | 0.0017b-f | 22.05bcd | 3.34d | 18.71abc | 2101.03abc |
| U-7 | Niaw dam | 25.79a-f | 44.06a-f | 18.51a-f | 0.0031a-f | 32.51abc | 6.94bcd | 25.56abc | 3250.59a |
| U-8 | Niew san pah tawng | 32.97abc | 85.43abc | 19.90a-d | 0.0053abc | 28.54abc | 7.84bcd | 20.70abc | 2853.59ab |
| U-9 | Khi Tom Gamnan | 28.15a-e | 90.33ab | 27.08a | 0.0054ab | 25.15abc | 9.68ab | 15.46bcd | 2514.62abc |
| U-10 | RD6 | 5.61g | 8.06ef | 4.61e-h | 0.0019b-f | 31.82abc | 7.91bcd | 23.91abc | 3181.57a |
| U-11 | Chum Phae 60 | 43.95a | 95.61a | 21.61abc | 0.0051a-d | 32.79abc | 8.28bcd | 24.51abc | 3279.41a |
| U-12 | Mahk Nam | 16.56b-g | 20.16c-f | 10.53b-h | 0.0033a-f | 33.02abc | 7.28bcd | 25.74ab | 3302.11a |
| U-13 | RD14 | 9.04efg | 27.36b-f | 18.91a-e | 0.0040a-f | 33.34abc | 6.37bcd | 26.97ab | 3334.42a |
| U-14 | RD-Maijo 2 | 33.73ab | 39.11a-f | 24.37ab | 0.0055ab | 21.40bcd | 6.58bcd | 14.81bcd | 2003.45abc |
| U-15 | KDML105 | 3.13g | 9.09ef | 4.40e-h | 0.0006f | 34.01abc | 5.47bcd | 28.54ab | 3401.29a |
| U-16 | Hawm | 3.13g | 6.16ef | 4.46e-h | 0.0005f | 28.86abc | 8.12bcd | 20.74abc | 2886.43ab |
| U-17 | RD8 | 10.26efg | 18.94c-f | 8.68c-h | 0.0010def | 33.12abc | 8.33bcd | 24.79abc | 3311.61a |
| U-18 | Khao Pahn Luang | 17.77b-g | 80.50a-d | 25.40a | 0.0050a-e | 31.82abc | 9.08abc | 22.74abc | 3071.14ab |
| U-19 | Plawng Aew | 7.12fg | 24.89b-f | 13.38a-h | 0.0020b-f | 35.25ab | 6.53bcd | 28.72ab | 3524.92a |
| U-20 | Pinkaset 3 | 31.98abc | 34.02a-f | 9.51c-h | 0.0018b-f | 21.59bcd | 5.87bcd | 15.71bcd | 2081.01abc |
| U-21 | Pahn Tawng | 2.13g | 8.07ef | 5.76d-h | 0.0011def | 24.26abc | 5.62bcd | 18.64abc | 2426.01abc |
| U-22 | Chai Nat 1 | 2.95g | 7.35ef | 4.61e-h | 0.0008f | 25.71abc | 7.39bcd | 18.32abc | 2571.23ab |
| U-23 | Hahng Yi 71 | 5.80fg | 17.24c-f | 12.51a-h | 0.0012 c-f | 31.28abc | 6.38bcd | 24.90abc | 2814.81ab |
| U-24 | TS2 | 3.29g | 5.04ef | 1.75h | 0.0004f | 25.21abc | 7.71bcd | 17.50abc | 2445.54abc |
| U-25 | Homwarin | 3.85g | 9.51ef | 5.65d-h | 0.0009ef | 29.75abc | 4.99bcd | 24.76abc | 2974.86ab |
| U-26 | Gai noi leuang | 20.72b-g | 37.46a-f | 13.17a-h | 0.0029a-f | 28.21abc | 4.57cd | 23.64abc | 1305.74bc |
| U-27 | Muey Nawng 62 M | 20.50b-g | 74.18a-e | 20.97abc | 0.0066a | 31.75abc | 13.05a | 18.70abc | 2833.22ab |
| U-28 | Goo Meuang Luang | 13.51c-g | 26.77b-f | 17.50a-g | 0.0013c-f | 31.36abc | 7.98bcd | 23.37abc | 3135.63a |
| U-29 | Beu PaTaw | 30.73a-d | 68.06a-f | 13.40a-h | 0.0020b-f | 9.06d | 6.26bcd | 2.80d | 815.02c |
| U-30 | Beu Saw Mi | 4.24g | 7.82ef | 4.89e-h | 0.0011def | 23.00bcd | 7.14bcd | 15.86bcd | 1980.81abc |
| U-31 | Hawm Bao Rai | 3.78g | 22.24c-f | 20.94abc | 0.0016 b-f | 36.05ab | 7.08bcd | 28.97ab | 3493.63a |
| P value | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | |
| CV (%) | 17.47 | 11.98 | 20.69 | 29.63 | 10.16 | 11.85 | 13.71 | 11.64 |
Note: Means in the same column followed by the same letters are not significantly different by DMRT. MS = mesocotyl length (mm), SL = shoot length (mm), COD = Coleoptile length of the deep-sowing experiment (mm), SW = shoot weight (g), COA = Coleoptile length of the anaerobic germination experiment (mm), CON = Coleoptile length of the normal condition (mm), AI = anaerobic response index, SV = seedling vigor.
associated with enhanced deep sowing tolerance. Mesocotyl elongation usually supports deep sowing. It is influenced by multiple genetic and environmental factors (Sakhale et al., 2023; Zhao et al., 2018).
In a separate selection index, which focused on traits related to anaerobic germination such as COA, CON, AI, and SV, five genotypes were identified: U-5 (Pin Gaew 56), U-19 (Plawng Aew), U-31 (Hawm Bao Rai), U-13 (RD14), and U-1 (RD45) (Fig. 4b). Notably, genotypes U-5 and U-1 are floating and deep-water rice cultivars that possess high individual genetic value. This finding is consistent with the observations of Suaythaisong et al. (2019), who reported that deep-water rice varieties exhibit adaptability to anaerobic germination conditions. This adaptability is linked to a high level of CIPK15 gene expression. This promotes starch mobilization and enhances coleoptile elongation, allowing the seedling to reach above the water surface and absorb oxygen. When constructing the selection index, all parameters from the evaluation were considered. Among the thirty-one varieties in this study, five varieties were selected: U-19 (Plawng Aew), U-11 (Chum Phae 60), U-31 (Hawm Bao Rai), U-5 (Pin Gaew 56), and U-2 (Prachinburi 2) (Fig. 4c). These varieties represent a new source of valuable materials for developing specific rice varieties with carried two important traits for direct-seeding rice production.
Materials and Methods
Plant materials and growing conditions
Thirty-one rice cultivars were collected for this study (Supplementary table 1) classified into three major ecotypes (lowland, upland, and floating/deep-water). These cultivars were transplanted 30 days after sowing in the experimental field at Faculty of Agriculture, Ubon Ratchathani University. A total of forty-eight uniform plants from each cultivar were transplanted in rows with a spacing of 25 × 25 cm². Fertilizer was applied at a rate of 75-50-75 kg N-P₂O₅-K₂O ha⁻¹, with applications at the seedling, tillering, and booting stages. Thirty days after heading, the yield was harvested and dried in an oven at 50 °C for four days to break dormancy.
Phenotyping for early seedling vigor
The experiments entered on evaluating two specific traits: deep-sowing tolerance and anaerobic germination tolerance. Aerobic germination conditions, which represented standard growth conditions, were used as the control. The evaluation experiments were replicated three times biologically for each rice varieties.
Evaluation of deep-sowing tolerance in rice
This experiment (adapted from Edzesi et al., 2023), aimed to measure mesocotyl length as an indicator of deep-sowing
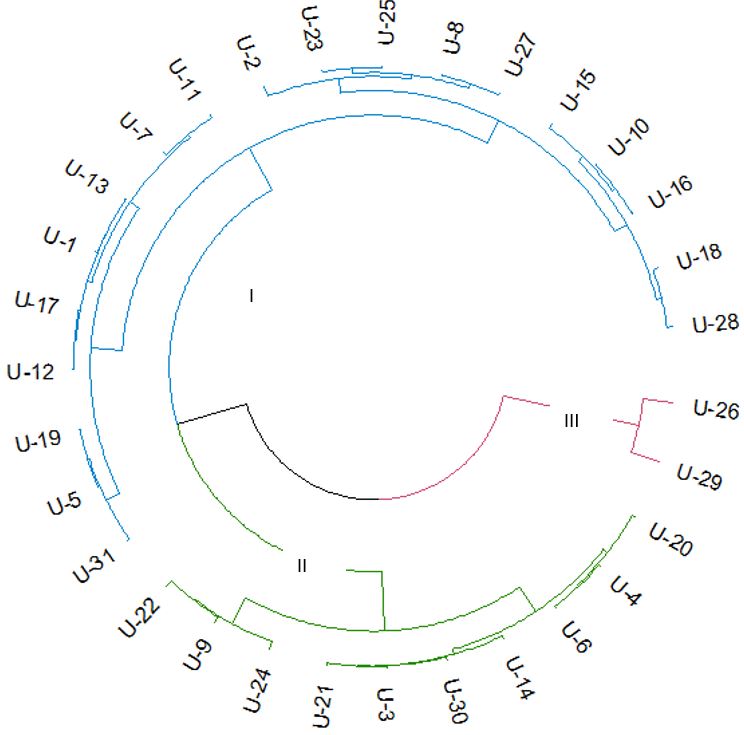
Fig 3. The hierarchical clustering dendrogram illustrates the grouping of 31 rice varieties based on traits associated with early seedling vigor parameters. The varieties are distributed into three clusters. Different colors were used to differentiate these clusters: blue for Cluster I, green for Cluster II, and red for Cluster III.
Table 3. Principal component analysis of related traits associated with early seedling vigor parameters in 31 rice varieties.
| Parameters | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 |
|---|---|---|---|---|---|---|---|---|
| MS | -0.4469 | 0.0935 | -0.3699 | -0.6602 | -0.0762 | 0.2453 | 0.3909 | -0.0039 |
| SL | -0.5041 | 0.0104 | -0.0481 | -0.1497 | -0.2419 | -0.6026 | -0.5463 | -0.0300 |
| COD | -0.4679 | -0.0995 | -0.1547 | 0.6727 | -0.4125 | 0.0914 | 0.3410 | 0.0011 |
| SW | -0.4887 | -0.0885 | -0.0114 | 0.1928 | 0.7366 | 0.3121 | -0.2745 | 0.0280 |
| COA | 0.0310 | -0.5773 | -0.0703 | -0.0443 | 0.1859 | -0.2650 | 0.2278 | -0.7086 |
| CON | -0.2723 | -0.1961 | 0.8465 | -0.1697 | -0.0042 | -0.0860 | 0.3052 | 0.2037 |
| AI | 0.1106 | -0.5382 | -0.3337 | -0.0137 | 0.1587 | -0.2956 | 0.1396 | 0.6742 |
| SV | 0.0336 | -0.5586 | 0.0634 | -0.1451 | -0.4040 | 0.5531 | -0.4389 | 0.0055 |
| eig | 3.6153 | 2.9224 | 0.8422 | 0.2697 | 0.1605 | 0.1090 | 0.0799 | 0.0010 |
Note: MS = mesocotyl length (mm), SL = shoot length (mm), COD = Coleoptile length of the deep-sowing experiment (mm), SW = shoot weight (g), COA = Coleoptile length of the anaerobic germination experiment (mm), CON = Coleoptile length of the normal condition (mm), AI = anaerobic response index, SV = seedling vigor.
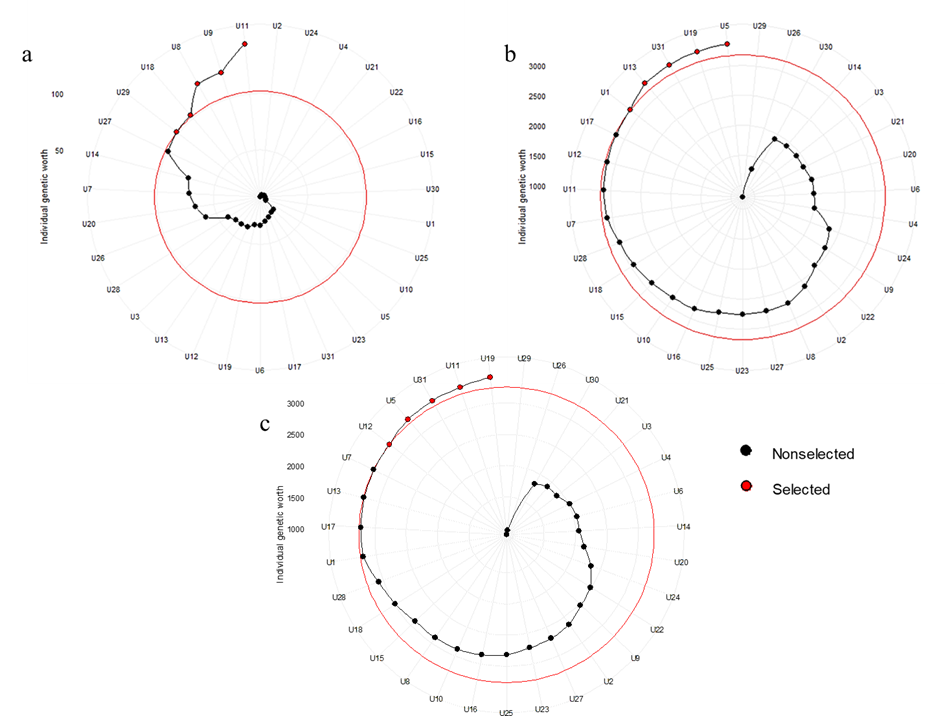
Fig. 4. The Smith-Hazel index, as demonstrated here, ranks rice varieties under a selection pressure of 20%. The index is based on combined parameters: (a) traits related to deep sowing, (b) traits related to anaerobic germination tolerance, and (c) a combination of all parameters. The black dots represent non-selected samples, while the red dots indicate selected ones. The values 1000, 1500, 2000, 2500, and 3000 represent the individual genetic worth. Out of the thirty-one varieties, five were selected under each condition as valuable genetic resources for enhancing direct-seeded rice varieties.
tolerance and early vigor-related traits. Thirty seeds were wrapped in 20 × 20 cm pieces of wet absorbent filter paper and placed vertically in a plastic container at 30 °C in complete darkness. Tap water was sprinkled to maintain moisture. Measurements of mesocotyl length (MS), seedling length (SL), coleoptile elongation (COD), and seed weight (SW) were taken after ten days using ImageJ.
Evaluation of anaerobic germination tolerance in rice
This experiment adapted from the methods of Hsu and Tung (2015) and comprised of placing 30 healthy seeds on water-soaked filter paper within a 12-cm tall plastic cup for the control treatment. In the submergence experiment, 30 healthy seeds were immersed in sterilized water to a depth of 10 cm within a 12-cm plastic cup, and maintained at 29–30 °C. After 7 days, coleoptile length was measured with ImageJ, and the anaerobic response index (AI) and seedling vigor (SV) were calculated using the following formulas:\(\ \)
\(AI = \ (submerged\ coleoptile\ length\ - \ control\ coleoptile\ length\))
\(SV = \ (Final\ germination\ percentage\ \times \ seedling\ length\))
Data analysis
Analysis of variance (ANOVA) and the 1% Duncan multiple range test (DMRT) were conducted using STAR IRRI (http://bbi.irri.org/products). Correlation analysis was performed with the Performance Analytics software package (https://cran.r-project.org/web/packages/PerformanceAnalytics/PerformanceAnalytics.pdf). Principal component analysis was carried out using the factoextra package (https://cran.r-project.org/web/packages/factoextra/index.html). A hierarchical clustering dendrogram was generated with the dendextend and circlize packages (https://r-charts.com/part-whole/circular-dendrogram). The Smith–Hazel index was analyzed using the Metan package for multi-environment trial analysis (https://tiagoolivoto.github.io/metan/reference/plot.sh.html) within the R program (https://cran.r-project.org/).
Conclusion
In summary, thirty-one rice varieties were evaluated to identify traits associated with early seedling vigor during direct seeding. The deep-sowing tolerance and anaerobic germination tests indicated that high heritability was observed for all the assessed agronomic traits. Coleoptile length under normal conditions and coleoptile length in deep-sowing experiments were identified as key traits for early vigor in direct-seeding seedlings. The genotypes U-19 (Plawng Aew), U-11 (Chum Phae 60), U-31 (Hawm Bao Rai), U-5 (Pin Gaew 56), and U-2 (Prachinburi 2) were selected as promising candidates for novel genetic resources aimed at improving direct-seeding rice varieties through a multi - trait selection index.
Acknowledgements
This research was funded by the Indigenous Food Research and Industrial Development Centre and Faculty of Agriculture, Ubon Ratchathani University.
Statement of contributions
Conceptualization, S.K. and S.W.; methodology; A.T. and S.K.; data curation, A.T. and S.K.; funding acquisition, S.K; writing—original draft, A.T.; writing—review and editing, S.K., A.T and S.W. All authors have read and agreed to the published version of the manuscript.
References
Angaji SA, Septiningsih EM, Mackill DJ, Ismail AM (2010) QTLs associated with tolerance of flooding during germination in rice (Oryza sativa L.). Euphytica. 172(2):159–168.
Anshori MF, Purwoko BS, Dewi IS, Willy S (2019) Selection index based on multivariate analysis for selecting doubled-haploid rice lines in lowland saline prone area. SABRAO J Breed Genet. 51(2):161-174.
Ashfaq M, Zhu R, Ali M, Xu Z, Rasheed A, Jamil M, Shakir A, Wu X (2023) Adaptation and high yield performance of honglian type hybrid rice in Pakistan with desirable agricultural traits. Agriculture. 13(2):242.
Barik J, Kumar V, Lenka SK, Panda D (2019) Genetic potentiality of lowland indigenous indica rice (Oryza sativa L.) landraces to anaerobic germination potential. Plant Physiol. Rep. 24(2):249–261.
Castano-Duque L, Ghosal S, Quilloy FA, Mitchell-Olds T, Dixit S (2021) An epigenetic pathway in rice connects genetic variation to anaerobic germination and seedling establishment. Plant Physiol. 186(2):1042-1059
Candido W, Silva CM, Costa ML, Almeida Silva BE, Miranda BL, Pinto JFN, Reis EF (2020) Selection indexes in the simultaneous increment of yield components in topcross hybrids of green maize. Pesq. agropec. bras., Brasília. 55: 1-8.6
Edzesi WM, Dang X, Liu E, Bandoh WKN, Gakpetor PM, Ofori DA, Hong D (2023) Screening germplasm and detecting QTLs for mesocotyl elongation trait in rice (Oryza sativa L.) by association mapping. BMC Genomics. 24(1):8.
Ghosal S, Casal C, Quilloy FA, Septiningsih EM, Mendioro MS, Dixit S (2019) Deciphering genetics underlying stable anaerobic germination in rice: phenotyping, QTL identification, and interaction analysis. Rice. 12(1):50.
Hossain MdM, Azad MAK, Alam MS, Eaton TEJ (2021) Estimation of variability, heritability and genetic advance for phenological, physiological and yield contributing attributes in wheat genotypes under heat stress condition. AM. J. Plant Sci. 12(04):586–602.
Hsu SK, Tung CW (2015) Genetic mapping of anaerobic germination-associated QTLs controlling coleoptile elongation in rice. Rice. 8(1):1–12.
Hu Z, Yan H, Yang J, Yamaguchi S, Maekawa M, Takamure I, Tsutsumi N, Kyozuka J, Nakazono M (2010) Strigolactones negatively regulate mesocotyl elongation in rice during germination and growth in darkness. Plant Cell Physiol. 51(7):1136–1142.
Ismail AM, Johnson DE, Ella ES, Vergara GV, Baltazar AM (2012) Adaptation to flooding during emergence and seedling growth in rice and weeds, and implications for crop establishment. AoB Plants. 2012:1-18.
Jahufer MZ, Casler MD (2015) Application of the Smith-Hazel selection index for improving biomass yield and quality of switchgrass. Crop Sci. 55(3):1212–1222.
Jang SG, Park SY, Lar SM, Zhang H, Lee AR, Cao FY, Seo J, Ham TH, Lee J, Kwon SW (2021) Genome-wide association study (GWAS) of mesocotyl length for direct seeding in rice. Agron. 11(12):2527.
Lee HS, Sasaki K, Higashitani A, Ahn SN, Sato T (2012) Mapping and characterization of quantitative trait loci for mesocotyl elongation in rice (Oryza sativa L.). Rice. 5(1):1–10.
Lee HS, Sasaki K, Kang JW, Sato T, Song WY, Ahn SN (2017) Mesocotyl elongation is essential for seedling emergence under deep-seeding condition in rice. Rice. 10(1):32
Lu Q, Zhang M, Niu X, Wang C, Xu Q, Feng Y, Wang S, Yuan X, Yu H, Wang Y, Wei X (2016) Uncovering novel loci for mesocotyl elongation and shoot length in indica rice through genome-wide association mapping. Planta. 243(3):645–657.
Mahender A, Anandan A, Pradhan SK (2015) Early seedling vigor, an imperative trait for direct-seeded rice: an overview on physio-morphological parameters and molecular markers. Planta. 241(5):1027–1050.
Mar Htwe N, Aye M and Nyein Thu C (2020) Selection index for yield and yield contributing traits in improved rice genotypes. IJERD. 11(2):86-91.
Nghi KN, Tagliani A, Mariotti L, Weits DA, Perata P, Pucciariello C (2021) Auxin is required for the long coleoptile trait in japonica rice under submergence. New Phytol. 229(1):85–93.
Nghi KN, Tondelli A, Valè G, Tagliani A, Marè C, Perata P, Pucciariello C (2019) Dissection of coleoptile elongation in japonica rice under submergence through integrated genome-wide association mapping and transcriptional analyses. Plant Cell Environ. 42(6):1832–1846.
Ramos Guimarães PH, Guimarães Santos Melo P, Centeno Cordeiro AC, Pereira Torga P, Nakano Rangel PH, Pereira de Castro A (2021) Index selection can improve the selection efficiency in a rice recurrent selection population. Euphytica. 217(5):95.
Sakhale S, Yadav S, Clark L, Lipka A, Kumar A, Sacks E (2023). Genome-wide association analysis for emergence of deeply sown rice (Oryza sativa) reveals novel aus-specific phytohormone candidate genes that conferred adaption to dry-direct seeding in the field. Front. Plant Sci. 14:1172816.
Shin NH, Han JH, Jang S, Song K, Koh HJ, Lee JH, Yoo S, Chin JH (2020) Early vigor of a pyramiding line containing two quantitative trait loci, phosphorus uptake 1 (Pup1) and anaerobic germination 1 (AG1) in rice (O. sativa L.). Agriculture. 10(10):1–11.
Simon A, Yuri S, Hironobu S, Kenji I (2012) Genotypic variation in coleoptile or mesocotyl lengths of upland rice (Oryza sativa L.) and seedling emergence in deep sowing. Afr. J. Agric. Res. 7(47):6239–6248.
Smiderle ÉC, Furtini IV, C CS, da Silva Botelho FBS, Resende MPM, Botelho RTC, Filho JMC, Castro AP, Utumi MM (2019) Index selection for multiple traits in upland rice progenies. Rev. Ciênc. Agrár. 42(1):1-10.
Su L, Yang J, Li D, Peng Z, Xia A, Yang M, Luo L, Huang C, Wang J, Wang H, Chen Z, Guo T (2021) Dynamic genome-wide association analysis and identification of candidate genes involved in anaerobic germination tolerance in rice. Rice. 14(1):1.
Toledo AMU, Ignacio JCI, Casal C, Gonzaga ZJ, Mendioro MS, Septiningsih EM (2015) Development of improved ciherang-sub1 having tolerance to anaerobic germination conditions. Plant Breed. Biotech. 3(2):77–87.
Uzair M, Patil SB, Zhang H, Kumar A, Mkumbwa H, Zafar SA, Chun Y, Fang J, Zhao J, Khan MR, Yuan S, Li X (2022) Screening direct seeding-related traits by using an improved mesocotyl elongation assay and association between seedling and maturity traits in rice. Agron. 12(4):975
Wang X, Xu Y, Li P, Liu M, Xu C, Hu Z (2019) Efficiency of linear selection index in predicting rice hybrid performance. Mol Breeding. 39(6):77.
Wang Y, Liu J, Meng Y, Liu H, Liu C Ye G (2021) Rapid identification of QTL for mesocotyl length in rice through combining QTL-seq and genome-wide association analysis. Front. Genet. 12:1–14.
Wu J, Feng F, Lian X, Teng X, Wei H, Yu H, Xie W, Yan M, Fan P, Li Y, Ma X, Liu H, Yu S, Wang G, Zhou F, Luo L, Mei H (2015) Genome-wide association study (GWAS) of mesocotyl elongation based on re-sequencing approach in rice. BMC Plant Biol. 15(1):1–10.
Xiong Q, Ma B, Lu X, Huang YH, He SJ, Yang C, Yin CC, Zhao H, Zhou Y, Zhang WK, Wang WS, Li ZK, Chen SY, Zhanga JS (2017) Ethylene-inhibited jasmonic acid biosynthesis promotes mesocotyl/coleoptile elongation of etiolated rice seedlings. Plant Cell. 29(5):1053–1072.
Yang B, Zeng J, Chen S, Li S, Wu L, Wan X (2022) Genome-wide association study reveals the genetic basis of seed germination in japonica rice. Agriculture. 13(1):118.
Zhang X, Lai Y, Meng Y, Tang A, Dong W, Liu Y, Liu K, Wang L, Yang X, Wang W, Ding G, Jiang H, Ren Y, Jiang S (2023) Analyses and identifications of quantitative trait loci and candidate genes controlling mesocotyl elongation in rice. J. Integr. Agric. 22(2):325–340.
Zhan J, Lu X, Liu H, Zhao Q, Ye G (2020) Mesocotyl elongation, an essential trait for dry-seeded rice (Oryza sativa L.): a review of physiological and genetic basis. Planta. 251(1):1–14.
Zhao Y, Zhao W, Jiang C, Wang X, Xiong H, Todorovska EG, Yin Z, Chen Y, Wang X, Xie J, Pan Y, Rashid Mar, Zhang H, Li J, Li Z (2018) Genetic architecture and candidate genes for deep-sowing tolerance in rice revealed by non-syn GWAS. Front. Plant Sci. 9:332.