
Aust J Crop Sci. 19(02):175-179 (2025) | ISSN:1835-2707
https://doi.org/10.21475/ajcs.25.19.02.p247
Effects of water and temperature on the physiological potential of Albizia polycephala (benth) killip seeds
Ivanildo Claudino da Silva, Luan Danilo Ferreira de Andrade Melo*, João Luciano de Andrade Melo Junior, Larice Bruna Ferreira Soares, Reinaldo de Alencar Paes, Thaíse dos Santos Berto, Keven Willian Sarmento Galdino da Silva, Natália Marinho Silva Crisóstomo, Adriana Guimarães Duarte, Jaqueline Figueredo de Oliveira Costa and Adrielle Naiana Ribeiro Soares Tenório
Federal University of Alagoas (UFAL), Agronomy Department, Campus of Engineering and Agrarian Sciences (CECA), BR 104/Norte - Km 85, Rio Largo, Alagoas, Brazil
*Corresponding author: Luan Danilo Ferreira de Andrade
Melo 
ORCID: https://orcid.org/0000-0002-3719-2873
Abstract: The significant presence of Albizia polycephala (Benth) Killip in various forest ecosystems highlights its potential to play a crucial role in biodiversity conservation and ecosystem stability. Understanding and promoting the use of this species can greatly contribute to the preservation and restoration of natural environments, as well as sustainable agricultural practices. Seed germination is a complex process influenced by several abiotic factors, notably moisture and temperature. These factors are critical, and under extreme conditions, they can cause irreparable damage to seeds. Therefore, this study aimed to assess the impact of varying water volumes in the substrate and temperatures on the physiological viability of A. polycephala seeds. The interaction between ecology and agriculture is essential to promote sustainable agricultural practices that respect natural ecosystems. Proper forest management can ensure biodiversity conservation and agroecosystem stability. Investing in agroforestry systems can contribute to the restoration of degraded landscapes, fostering harmonious coexistence between agricultural production and environmental preservation.The experiment was conducted at the Fitotechnology Laboratory, Campus of Engineering and Agricultural Sciences, Federal University of Alagoas (CECA-UFAL), in Rio Largo – AL. Treatments included water volumes equivalent to 1.5, 2.0, 2.5, 3.0, and 3.5 times the dry mass of the substrate, with constant temperatures of 30°C and alternating temperatures of 20-30°C. Substrates were not rewetted during the experiment. A completely randomized design (CRD) was employed, and when significance was observed in the F-test, means were compared using Tukey's test at a 5% probability level. Additionally, regression analysis was conducted to analyze quantitative effects. The alternating temperature regime of 20-30°C and a water volume of 3.5 times the mass of the dry substrate proved to be the most favorable combination for conducting germination and vigor tests of A. polycephala seeds. These findings can inform ecological restoration projects and agricultural strategies aimed at enhancing seed viability and supporting ecosystem resilience.
Keywords: Agroecosystem, Ecological impact, Restoration of degraded areas.
Abbreviations: BOD_Biochemical Oxygen Demand; IVG_Index of speed of germination; UFAL_Federal University of Alagoas; CRD_completely randomized design.
Introduction
Albizia polycephala (Benth) Killip, popularly known as angico-branco, belongs to the Fabaceae – Mimosoideae family and is a pioneer species with multiple uses, including environmental restoration, ornamentation, and urban afforestation (Silva et al., 2023). Species of the genus Albizia, particularly A. polycephala, are widely recognized for their potential in forestry and agroforestry practices. They are considered excellent options for the restoration of successional areas and forest formations, playing a crucial role in natural ecosystems, such as the Atlantic Forest, where they exhibit high frequency, density, and relative dominance (Sartori et al., 2018). Their presence in various forest types underscores their ability to maintain biodiversity and stabilize agroecosystems (Melo et al., 2018). Therefore, understanding and promoting the use of native species can significantly contribute to the conservation and restoration of natural environments (Melo et al., 2021).
Water is a fundamental solvent for biological processes, essential for the movement of molecules within and between cells, promoting metabolism. In the context of seeds, water is crucial for the germination process, activating their metabolic system through physiological and biochemical changes (Paiva et al., 2019). This activation results in cell division and expansion, enabling the growth of the embryo and the protrusion of the primary root (Gonçalves et al., 2015). For this metabolic activation to occur, seeds need to reach an appropriate level of hydration. However, excess water can be detrimental to the germination process (Taiz et al., 2017). Understanding the optimal water conditions in germination tests ensures greater seed trade security through standardized analyses for evaluating different seed lots (Brasil, 2009; Leão et al., 2019). Excessively waterlogged substrates reduce aeration, hinder oxygen penetration, and decrease metabolic processes, leading to a reduction in the physiological potential of seeds. Additionally, excess water can increase fungal incidence, contributing to seed deterioration (Melo et al., 2017). Therefore, during germination tests, it is crucial to maintain the substrate uniformly moist to provide seeds with the necessary amount of water for proper germination and development, ensuring favorable conditions for germination success and seedling establishment (Oliveira et al., 2022; Silva et al., 2024).
Temperature plays a key role in seedling and plant development, influencing cellular metabolic reactions and significantly impacting physiological activities in both natural environments and controlled cultivation conditions (Melo Junior et al., 2018; Araújo et al., 2014). Research by Melo et al. (2021) investigates the effects of different temperature ranges on the development of cultivated forest species, aiming to optimize seed germination. The optimal temperature range is the one that allows the highest percentage of germination in the shortest period of time (Crisostómo et al., 2021). Managing temperature during the germination process can be an efficient strategy to maximize the percentage, speed, and uniformity of germination, resulting in more vigorous plants and increased efficiency in seedling production (Melo et al., 2021). The temperature during the seed hydration phase plays a critical role, influencing the viscosity of water and, consequently, the water flow from the environment to the seed, directly affecting the metabolic reactions associated with the germination process (Leão et al., 2019).
Based on this, the objective of the present work was to evaluate the influence of different volumes of water in the substrate and temperatures on the physiological potential of A. polycephala by understanding these ecological factors, we aim to enhance the effectiveness of seed germination and contribute to both ecological restoration efforts and sustainable agricultural practices.
Results and Discussion
Seed moisture content and vigor
At the time of harvest, the moisture content of Albizia polycephala seeds was 12.4%. Analyzing the results regarding seed vigor defined by the first germination count (Figure 1), it can be seen that seed vigor was higher (52%) when using a volume of 3.5 times the mass of dry paper at an alternating temperature of 20-30°C, which differed statistically from the constant temperature of 30°C. At a temperature of 30°C, the maximum percentage (45%) was also recorded when the substrate was moistened with a volume of water equivalent to 3.5 times the weight of the dry substrate. Previous studies have found similar results across different types of seeds. For example, Araújo et al. (2014) observed that Luffa operculata seeds had a higher germination rate in the first count when exposed to temperatures of 20-30°C and 35°C, and subjected to volumes of water equivalent to 2.3 and 3.5 times the weight of the dry paper, respectively. Similarly, Oliveira Junior et al. (2015) discovered that seeds of Hylocereus costaricensis had a higher germination rate at the first count when subjected to a temperature of 30°C and water volumes exceeding 3.0 times the weight of the dry paper. Additionally, José et al. (2011), also supporting the present study, found that moistening at 3.0 times the mass of the dry paper yielded good results in the first germination count for Apuleia leiocarpa.
Effects of substrate moistening levels
Melo (2017) reported that higher levels of substrate moistening when sowing between paper had a positive effect on the germination performance of Mimosa bimucronata seeds. Conversely, Martins et al. (2009) studied substrate moistening levels on the germination of Bactris gasipae seeds and found a deleterious effect due to increasing water levels. Such studies are essential, as the amount of water in the substrate that enables good germination performance of seeds depends on the species' tolerance to extreme water conditions—whether excess or lack—due to ecological adaptation to their place of origin (Silva et al., 2017). Lower germination performance was observed in substrates moistened with less than 2.0 times the mass of dry paper. At moisture levels of 2.0 and 1.5 mL/g right on the date of the first count, the substrate had already dehydrated after four
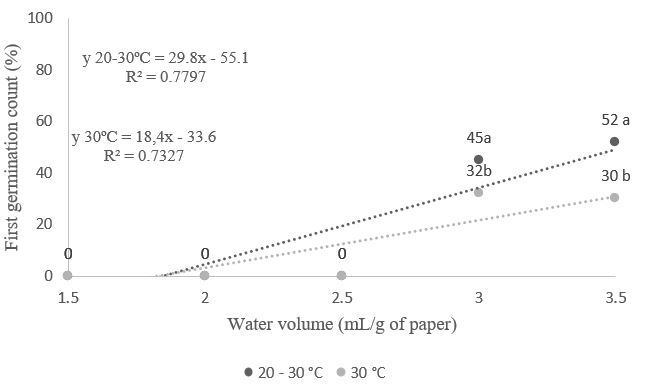
Figure 1. First germination count (%) of Albizia polycephala (Benth.) Killip ex Record seeds, subjected to different temperatures and volumes of water in the substrate (CECA/UFAL, 2024).
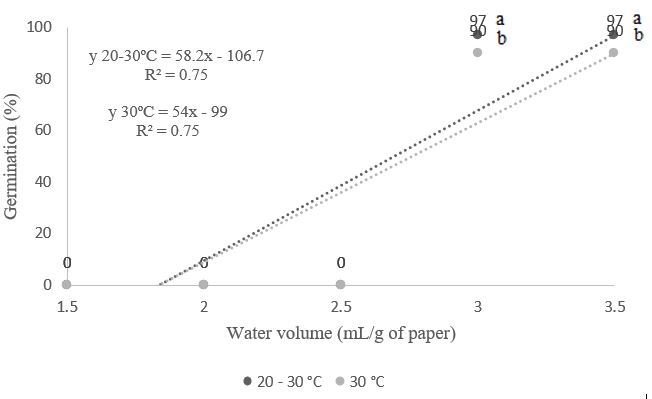
Figure 2. Germination percentage (%) of Albizia polycephala (Benth.) Killip ex Record seeds, subjected to different temperatures and volumes of water in the substrate.
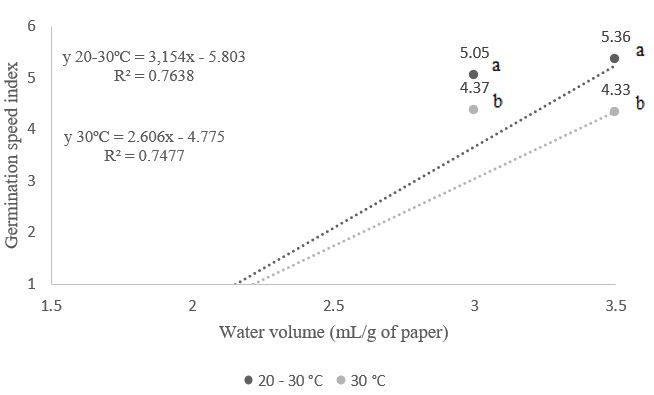
Figure 3. Germination speed index of Albizia polycephala (Benth.) Killip ex Record seeds, subjected to different temperatures and volumes of water in the substrate.
days, making it difficult to continue the test. Thus, these moisture levels are not suitable for routine work. Neves (2021), in research with different volumes of water in the substrate, concluded that moistening levels below 2.0 times its weight in water hindered handling and were not suitable for routine work in a seed analysis laboratory, confirming the findings of this research. Padilha et al. (2018) used the first germination count as a method to evaluate differences in the amount of water in the Apuleia leiocarpa seed germination test. In this sense, the substrate between germitest paper favors the expression of vigor in Albizia polycephala seeds, possibly related to the greater contact surface that the substrate provides.
Impact of water volume on germination speed index and seedling growth
Germination percentages were higher when a water volume of 3.5 times the mass of the dry substrate was used at both temperatures (30°C and 20-30°C), statistically differing from each other, reaching 90% and 97%, respectively (Figure 2). Gonçalves et al. (2015), studying Parkia platycephala seeds, found that using different water volumes (2.0, 2.5, 3.0, and 3.5 times the mass of dry paper) and temperatures (25, 30, and 20-30°C) enabled better expression of the physiological potential of the seeds. For Amburana cearensis seeds, a volume of water equivalent to 3.25 times the weight of the dry substrate at a temperature of 30°C provided a higher percentage of germination (Guedes et al., 2010). Despite the differences among species, the need for a greater amount of water in the substrate for the good development of A. polycephala is evident. High humidity conditions may lead to a lack of O2 during the germination process, reducing respiration, delaying, or even arresting the development of the embryonic axis, potentially increasing the occurrence of abnormal seedlings (Carvalho & Nakagawa, 2012); however, this did not occur in this research.
A linear increase in the germination speed index (GSI) was observed at temperatures of 30°C and 20-30°C (Figure 3), corresponding to the gradual increase in the volume of water used to moisten the paper, with statistical differences observed. This increase occurred from a volume of 2.0 times the mass of dry paper; lower water volumes did not allow for observations, as no seeds germinated due to the insufficient water supply. It is likely that the substrate used in this research provided less water to the seeds when below the volume of 2.0 times the mass of dry paper, prolonging phase II of the germination process, which depends on the substrate's water potential, thereby delaying germination. As reported by Padilha et al. (2018), lower water potential in the substrate extends this phase.
Alves & Cardoso (2020) found different results with Leucaena leucocephala, where the lowest germination speed index was observed when using a water proportion equivalent to 3.0 times the weight of the dry substrate. The hydration temperature can significantly alter seed viability and vigor. Water plays a critical role in the development process, as seeds transition from a metabolically active state to an inactive one after maturation due to desiccation, returning to a metabolically active state during germination (Rahman et al., 2011). Melo (2017) found, with Mimosa bimucronata seeds, a linear increase in GSI at temperatures of 30°C and 20-30°C, due to increased water volumes used to moisten the substrate, results consistent with this research.
The use of water volumes of 1.5 and 2.0 times the mass of the dry substrate (Figures 1, 2, 3, 4, and 5) did not yield any germinated seedlings at both temperatures studied, highlighting a more pronounced effect of temperature on germination as water levels were gradually raised. Generally, Taiz et al. (2017) noted that water deficit interferes not only with the germination process of seeds but also diminishes seedling development by reducing cellular pressure. Based on Bewley & Black (1994), the reduction in seedling dry mass due to water deficiency occurs from delays in biochemical and physiological processes or difficulties in hydrolysis and the mobilization of reserves stored in the seeds.
Regarding the growth and dry mass of A. polycephala seedlings, with a water volume of 3.5 times the mass of the dry substrate, seedlings achieved lengths of 13.2 cm and 14.0 cm at temperatures of 30°C and 20-30°C, respectively (Figure 4). For dry mass, seedlings reached 0.101 g and 0.132 g (3.5 times the mass of the dry substrate) at temperatures of 30°C and 20-30°C, respectively (Figure 5). Silva et al. (2018) and Oliveira et al. (2019) found that hydration temperature can significantly modify seed prediction and vigor; therefore, rehydration is considered essential in the development process (Duarte et al., 2018).
When the substrate was moistened below 2.0 mL/g, a negative impact on A. polycephala seeds was observed, affecting both germination and seedling growth regardless of the temperature used (Figures 1, 2, 3, 4, and 5). This effect can be attributed to
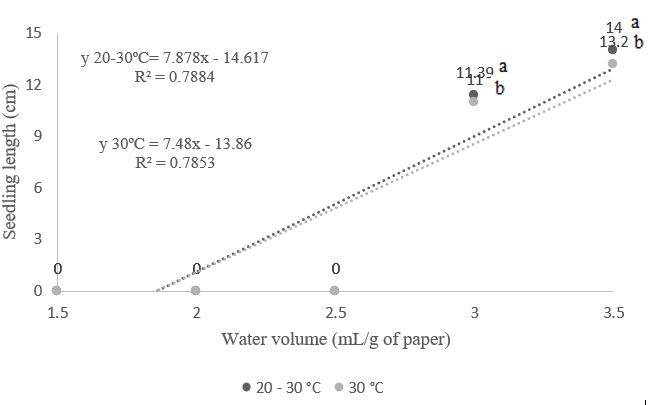
Figure 4. Length of seedlings from Albizia polycephala (Benth.) Killip ex Record seeds, subjected to different temperatures and volumes of water in the substrate.
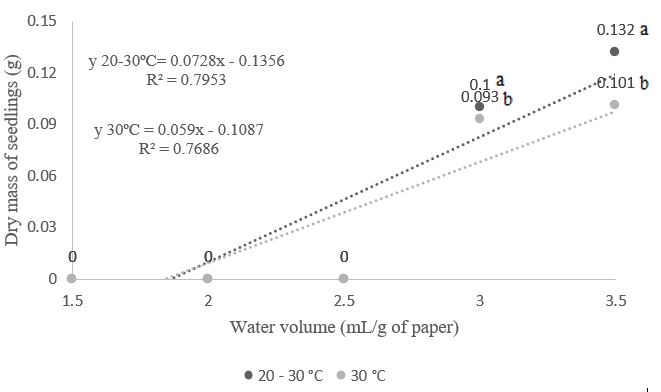
Figure 5. Dry mass of seedlings from Albizia polycephala (Benth.) Killip ex Record seeds, subjected to different temperatures and volumes of water in the substrate.
potential water restrictions due to inadequate water in the germination environment, leading to reduced respiration and, consequently, delays or interruptions in germination (Carvalho & Nakagawa, 2012; Marcos Filho, 2015). In studies with Amburana cearenses seeds, Guedes et al. (2010) found that a water volume of 3.5 times the mass of the dry substrate and a temperature of 30°C were optimal for seedling growth, resulting in higher dry mass content. The same trend was observed in this experiment for the alternating temperature of 20-30°C.
The present results align with those reported by Afonso et al. (2017), who, while analyzing Jacaranda copaia seeds, found that the greatest seedling lengths were achieved when seeds were germinated at temperatures of 25°C and 30°C, with no statistical differences in the amount of water or the interaction between factors. Working with Amburana cearensis seeds, Medeiros et al. (2020) observed that a water volume of 3.5 times the mass of the dry substrate led to better seedling growth, a result similar to what was found in this experiment.
The volume of water also influences the imbibition process. Although each species has unique characteristics, imbibition involves the process of water absorption by the seed, consisting of three phases, which provides an increase in the respiratory rate, ultimately leading to the rupture of the seed coat through the root axis. This process is crucial for initiating germination, as it activates metabolic pathways necessary for the development of the seedling. The efficient uptake of water during imbibition can significantly affect the timing and success of germination, highlighting the importance of optimal moisture levels for different seed species.
Materials and methods
Location and seed collection
The experiment was conducted at the Phytotechnics Laboratory of the Agricultural Engineering and Sciences Campus at the Federal University of Alagoas (CECA-UFAL), Rio Largo - AL, Brazil. Seeds of Albizia polycephala (Benth.) Killip ex Record were obtained from ripe fruits harvested from nine mother trees in Recife – PE (latitude 8º 04' 03''S, longitude 34º 55' 00''W). After harvesting, the fruits were manually processed by making a longitudinal cut in the pod to extract the seeds. These seeds were then used for germination and vigor tests.
Experimental design
The experimental design was completely randomized (CRD), with four replications of 25 seeds per treatment. Seeds were placed to germinate between two sheets of Germitest paper in transparent Gerbox® plastic boxes (SILVA et al., 2023). The paper was moistened with distilled water at volumes equivalent to 1.5, 2.0, 2.5, 3.0, and 3.5 times the dry weight of the paper, with no subsequent water addition (Melo, 2017). Germination occurred in a Biochemical Oxygen Demand (B.O.D.) chamber set at a constant temperature of 30°C and alternating temperatures of 20-30°C, with an 8-hour photoperiod. The Gerbox® containers were stored in transparent plastic bags to prevent moisture loss.
Germination evaluation
Evaluations were performed daily over 15 days, after which germination stabilized. Seeds were considered germinated if they produced a primary root, hypocotyl, and epicotyl, all free of defects and viable for field planting (Brasil, 2009).
Physiological quality assessments
Germination: Daily counts were made for 15 days, with seeds considered germinated when the primary root reached ≥ 2 mm (Silva et al., 2023).
First Germination Count: This was performed on the fourth day after sowing, providing an accumulated percentage of germinated seeds (Melo et al., 2018).
Germination Speed Index (GSI): Germination counts were conducted daily until stabilization, and the GSI was calculated using the formula proposed by Maguire (1962).
Seedling Length: At the end of the germination test, the shoot and root lengths of seedlings were measured with a ruler, and the results were expressed in centimeters per seedling (Melo et al., 2021).
Seedling Dry Mass: After the germination test, normal seedlings were dried in Kraft paper bags in a forced-air oven at 80°C for 24 hours. The dried samples were then placed in desiccators and weighed on an analytical balance with an accuracy of 0.0001g, with the results expressed in g/seedling (Nakawaga, 1999).
Statistical analysis
Where the F-test indicated significance, means were compared using Tukey's test at 5% probability. For quantitative effects, polynomial regression analysis was applied to assess linear, quadratic, and cubic effects of the variables. The model with the highest order and the greatest determination value was chosen to represent the behavior of each variable. Statistical analyses were performed using SISVAR 5.6 software from the Federal University of Lavras (Ferreira, 2014).
Conclusion
The temperature alternately of 20-30°C and the water volume of 3.5 times the mass of the dry substrate is the most suitable combination for carrying out germination and vigor tests on Albizia polycephala seeds.
References
Afonso MV, Martinazzo EG, Aumonde TZ, Villela FA (2017) Physiological parameters of Albizia niopoides seedlings produced in different substrate compositions. Ciência Florestal. 27(4): 1395–1400 (Br).
Alves AU, Cardoso EA (2020) Substrate moistening and temperature in leucaena seed germination. In: Engenharia Florestal: Desafios, Limites e Potencialidade, Científica Digital, São Paulo, Brasil, 868-878 (Br).
Araújo PC, Alves EU, Ursulino MM, Araújo LR, Silva RS (2014) Different water volumes in the substrate and temperatures for germination of cabacinha seeds. Revista Horticultura Brasileira. 32(3): 367-370.
Araujo MMV, Fernandes DÁ, Jardini DC, Camili EC (2019) Pre-hydration and physiological conditioning of yellow passion fruit seeds. Revista Agro@mbiente. 11(3): 241-247 (Br).
Barbosa LM, Shirasuna RT, Lima FC, Ortiz PRT (2015) List of species recommended for ecological restoration for different regions of the State of São Paulo. In: Simpósio de Restauração Ecológica. Anais Instituto de Botânica, São Paulo, 303-436 (Br).
Bewley JD, Black M (1994) Seeds: physiology of development and germination. 2. ed. Plenum Press, New York.
Brasil (2009) Ministry of Agriculture, Livestock, and Supply. Rules for Seed Analysis. Available at https://www.gov.br/agricultura/pt-br/assuntos/insumos-agropecuarios/arquivos-publicacoes-insumos/2946_regras_analise__sementes.pdf (Br).
Carvalho NM, Nakagawa J (2012) Seeds: science, technology and production. Funep, Jaboticabal, 524-590 (Br).
Carvalho CA, Aguiar EA, Teixeira DL, Delfino JS, Nascimento MM, Ferreira RF, Andrade RA, Brito RS (2020) Accelerated aging test to analyze the vigor of monkey ear seeds (Enterolobium schomburgkii Benth). Revista Thema, 17(2), 346-353 (Br).
Cipriani VB, Garlet J, Lima BM (2019) Breaking dormancy in Chloroleucon acacioides and Senna macranthera seeds. Revista de Ciências Agrárias, Lisboa, 21, 49- 54 (Br).
Crisostomo NMS, Santos LGV, Melo LDFA, Melo Junior JLA, Soares LBF, Paes RA, Oliveira VC, Chaves LFG, Costa JFO, Silva EE (2021) Seed germination of Enterolobium contortisiliquum (Vell.) Morong. (Fabaceae) under different temperatures and drying method. Journal of Scientific Research and Reports. 11: 54-59.
Duarte MM, Kratz D, Carvalho RLL, Nogueira AC (2018) Influence of water stress on seed germination and seedling formation of white angico. Advances in Forest Science. 5(3): 75-379 (Br).
Farias CCM, Lopes JC, Mengarda LHG, Maciel KS, Moraes CE (2019) Biometrics, physical characteristics and water absorption of Enterolobium maximum Ducke seeds. Ciência Florestal. 29(3): 1241- 1253 (Br).
Ferreira DF (2014) Sisvar: a Guide for its Bootstrap procedures in multiple comparisons. Ciência e Agrotecnologia. 38(2): 109-112.
Gomes JP, Oliveira LMD, Ferreira PI, Batista F (2016) Substrates and temperatures for germination testing on Myrtaceae seeds. Ciência Florestal. 26: 285-293 (Br)
Gonçalves EP, França PRCD, Viana JS, Alves EU, Guedes RS, Lima CRD (2015) Substrate moistening and temperature in the germination of Parkia platycephala Benth seeds. Ciência Florestal. 25: 563-569. (Br)
Guedes RS, Alves EU, Gonçalves EP, Viana JS, França PRCD, Lima CRD (2010) Substrate moistening and temperature on germination and vigor of Amburana cearensis (All) AC Smith seeds. Revista Brasileira de Sementes. 32: 116-122. (Br)
Inocente MC (2018) Water deficit in Eugenia spp seeds (Myrtaceae): resistance and relationships with the regenerability of roots and seedlings Dissertation, São Paulo, Brasil (Br).
José AC, Coutinho AB, Erasmo EAL (2011) Effect of temperature and substrate on the germination of Apuleia leiocarpa (Vogel) JF Macbr (amarelão) seeds. Agrarian: 4(14): 286-293.
Leão N, Campos MV, Felipe SH, Shimizu E (2019) Influence of the amount of water in the substrate on the germination of pau-preto (Cenostigma tocantinum Ducke) seeds Enciclopédia Biosfera. 16(29) (Br).
Maguire JD (1962) Speed of germination: aid in selection and evaluating for seedling emergence and vigour. Crop Science. 2(1): 176-177
Marcos Filho J (2015) Physiology of cultivated plant seeds FEALQ, Piracicaba (Br).
Martins CC, Bovi MLA, Spiering SH (2009) Substrate moistening in the emergence and vigor of peach palm seedlings. Revista Brasileira de Fruticultura. 31: 224-230. (Br)
Medeiros JADD, Nunes SPL, Félix FC, Ferrari CDS, Pacheco MV, Torres SB (2020) Vigor test of (strong) normal intact Amburana cearensis (Allemão) AC Smith seedlings. Journal of Seed Science. 42: e202042011
Melo Junior JLDA, Melo LDFDA, Ferreira VM, Araújo Neto JCD, Silva CBD, Neves MIRDS (2018) Thermal-biological aspects of seed germination of Colubrina glandulosa Perkins under different temperatures. Journal of Agricultural Science. 10(6): 390-400.
Melo LDFA (2017) Morphometry, physiological potential of seeds and post-seminal development of Mimosa bimucronata (DC) O KTZE Ph. D thesis Federal University of Alagoas, Engineering and Agricultural Sciences Campus, Brazil (Br)
Melo LDFA, Melo Junior JLA, Magalhaes ID, Medeiros AS, Maia Júnior SO, Cordeiro Junior JJF, da Silva AC (2017) Temperature and substrate effects on the germination of Caesalpinia ferrea Mart Ex Tul. African Journal of Agricultural Research. 12(47): 3348-3354.
Melo LDFA, Melo Junior JLA, Araujo Neto JC, Ferreira VM, Silva AC, Silva VSG (2018) Cardinal temperatures for the germination of Chorisia speciosa A St-Hil and parameters of the accelerated aging test for determination of vigor. Australian Journal of Crop Science. 12(10): 1653-1659.
Melo LDFA, Melo Junior JLA, Soares LBF, Chaves LFG, Araujo Neto JC, Ferreira VM, Neves MIRS, Goncalves EP, Viana JS, Costa JFO, Assis WO (2021) Thermal-biological aspects of germination of seeds in tropical forest tree species. Australian Journal of Crop Science. 15(6): 955-959.
Moraes CE, Lopes JC, Farias CCM, Maciel KS (2016) Physiological quality of Tabernaemontana fuchsiaefolia A DC seeds as a function of the accelerated aging test. Ciência Florestal. 26: 213-223 (Br).
Nakagawa J (1999) Seed vigor tests based on seedling performance In: Krzyzanowski, F C; Vieira, R D; França Neto, J B Seed vigor: concepts and tests Londrina: Informativo ABRATES. 21-224.
Neves BR (2021) Envelope method in the germination test of Panicum maximum Jacq. Ph, D thesis, São Paulo State University, Brazil (Br)
Oliveira Junior JL, Almeida Neta MN, David AMSS, Aguiar ACM, Gomes AGO, Amaro HTR, Donato LMS (2015) Substrate moistening and temperature on the germination and vigor of pitaya seeds. Comunicata Scientiae. 6(3): 282-290 (Br).
Oliveira SS, Nascimento GO, Souza DP, Nascimento LO, Oliveira SS, Gonçalves JFC, Ferreira Jb, Oliveira E (2019) Growth of parica seedlings (Schizolobium amazonicum Huber ex Ducke) cultivated in diferente organic substrates. Afr J Agric Res. 16: 303–310.
Oliveira RL, Didolanvi OD, Menezes ACP (2022) Substrate moistening and temperature in the germination of Anadenanthera macrocarpa seeds. Revista Ouricuri. 12(1): 1-10 (Br).
Padilha MS, Sobral LS, Baretta CRDM, de Abreu L (2018) Substrates and moisture content for the Apuleia leiocarpa (Vog) Macbr seed germination test. Revista Verde de Agroecologia e Desenvolvimento Sustentável. 13(4): 437-444 (Br).
Paiva EP, Torres SB, Morais JR, Sá FV, Leite MS (2019) Germination of chia seeds in diferente substrates and water volumes. Revista Caatinga. 12(1): 270-275
Rahman MM, Ahammad KU, Alam MM (2011) Effect of soaking condition and temperature on imbibition rate of maize and chickpea seeds. Research Journal of Seed Science. 4(2): 117-124.
Sartori RA, Balderi F, do Carmo TO (2018) Evaluation of Albizia polycephala (Benth) Killip germination in different substrates and light. Natureza Online. 16(2): 001-009 (Br).
Silva GZ, Bruno RDLA, Martins CC, Azevedo AIB, de Azevedo CF, Lima RS (2017) Morphoanatomy of Myracrodruon urundeuva Fr All seedlings submitted to different levels of water in the soil. Bioscience Journal. 33(5): 1321-1331
Silva AR, Leão-Araújo EF, Rezende BR, Santos WV, Santana HA, Silva SCM, Fernandes NA Costa DS, Mesquita JCP (2018) Modeling the three phases of the soaking kinetics of seeds. Agronomy Journal. 110(1): 164-170
Silva IC, Melo LDFA, Melo Junior JLA, Paes RA, Costa JFO, Oliveira VC, Duarte AG, Crisostomo NMS, Aureliano RHS, Silva JAC, Abreu LA (2023) Biometric characterization, post-seminal development and overcoming seed dormancy of Albizia polycephala (Benth) Killip ex Record. Australian Journal of Crop Science. 5: 524-530
Silva EE, Melo LDFA, Melo Junior JLA, Gomes LCA, Berto TS, Crisostomo NMS, Oliveira VC, Ramos MGC, Costa JFO, Soares ANR (2024) Biometry and physiological potential of Pterogyne nitens Tull seeds as a function of different substrates. Revista de Gestão e Secretariado. 15: 1-18.
Taiz L, Zeiger E, Møller IM, Murphy A (2017) Plant physiology and development 6 ed Artmed, Porto Alegre (Br).