
Aust J Crop Sci. 18(12):825-830 (2024) | ISSN:1835-2707
https://doi.org/10.21475/ajcs.24.18.12.p98
Identifying soybean progenies with high grain productivity and stress resilience to abiotic stresses
Willyan Júnior Adorian Bandeira1, Ivan Ricardo Carvalho1*, Murilo Vieira Loro2, José Antonio Gonzalez da Silva1, João Pedro Dalla Roza1, Victor Delino Barasuol Scarton1, Gabriel Mathias Weimer Bruinsma1, Leonardo Cesar Pradebon1
1Regional University of the Northwest of the State of Rio Grande do Sul, Ijuí, Rio Grande do Sul, Brazil
2Federal University of Santa Maria, Santa Maria, Rio Grande do Sul, Brazil
*Corresponding author: Ivan Ricardo Carvalho 
Abstract: This work aimed to select transgressive soybean progenies aiming at grain productivity and resilience to abiotic stresses. An incomplete block design augmented with interspersed controls was used, with segregating F2 populations conducted in four different environments. The number of seeds per plant (NSP) and seed weight per plant (SWP) were determined for all plants selected in the field, to determine the superiority of the genotypes in comparison with 25 commercial soybean controls. The REML (restricted maximum likelihood) methodology was used to estimate genetic parameters and variance components. Best Linear Unbiased Prediction (BLUP) was used to identify populations with transgressive genotypes for the evaluated traits. The Index of Genetic Selection for Resilience (IGSR∆) was also calculated for each selected genotype, as a way of determining the genetic gains of each population. Heritability values of 0.25 and 0.24 were observed for NSP and SWP, respectively. 264 superior lines were selected for SWP and 273 superior lines for NSP, with the largest number of superior genotypes for both variables selected in the environments of Boa Vista do Cadeado – RS and Eugênio de Castro – RS. The progenies selected from populations IRC 002, IRC 012, IRC 013, IRC 034 showed high potential. The progenies selected for IRC 032 and IRC 049 showed the greatest potential for resilience.
Keywords: Glycine max L., plant breeding, genotypes x environments, grain productivity, resilience, biometry.
Introduction
The grains produced by soybeans are sources of nutrients and proteins (Loro et al., 2021), used in the feed industry and for human consumption (Chen et al., 2023). Soybean production for Brazil in the 2022/23 harvest is estimated at 155 million metric tons, 16% higher than in the 21/22 harvest, with an expected average productivity of 3.55 tons per hectare and a cultivated area of 43.7 million hectares (USDA, 2023).
Due to their polygenic nature, grain yield and other quantitative traits are continuously controlled and affected by quantitative trait loci, genomic regions with associated genes, and environmental effects (Said et al., 2022). The evaluation of the genotype-environment interaction is important, as a genotype may be superior in certain environmental conditions, but may not present superiority in other environmental conditions, directly modifying the selection gains (Scarton et al., 2023). Studies of the genotype-environment interaction of soybean crops have been conducted, mainly using methods to evaluate adaptability and stability for agronomic (Almeida et al., 2021; Albuquerque et al., 2022; Pradebon et al., 2023) and nutritional traits of grains (Sangiovo et al., 2023).
The genetic gains in the breeding population in each selective cycle must be maximized for a condition of restriction on the effective population size (Resende and Alves, 2021), for which estimates of genetic parameters are fundamental in the genetic gain in each generation. Heritability, as an indication of genetic superiority that can be transmitted to progenies, is a good parameter for quantifying the advancement of selection. For this reason, progeny testing in different environments is used as it favors the selection of complex inheritance characteristics, determined by several genes and influenced by the environment (Laviola et al., 2014), where the quantification of the effect of the environment on variation residual is a key element for understanding phenotypic expression (Loro et al., 2022).
Strategies that can be used to obtain gains in a short period include choosing the best method to be used and the selection pressure that will be used (Facchinello et al., 2021). For this, genetic variability is essential for evolutions in the improvement of plant species (Singh et al., 2021), and the use of high selection pressures in initial generations reduces genetic variability in subsequent generations (Facchinello et al., 2021). Using methodologies based on linear mixed models is possible to include all sources of variation to estimate precise genetic values, without considering the effect of the environment. The methodology called Restricted Maximum Likelihood/Best Linear Unbiased Prediction (REML/BLUP) can be satisfactorily used to infer the genotype-environment interaction, which makes use of random effects to estimate the means of individuals in a population (Resende, 2007). In this context, the objective of this work was to select transgressive soybean progenies aiming at grain productivity and resilience.
Results and Discussion
Climatological information
The air temperature was mild in the months of October, November and April, with the average temperature varying between 18 ºC and 22 ºC, with a greater thermal amplitude in the months of October, November and April (Figure 1). The average temperature values were within the ideal range for vegetative development and flowering in all environments, remaining above the optimal temperature for grain filling in March. Precipitation volumes were below normal, with accumulations below 25 mm in all months, except for the environments of Ijuí and Eugênio de Castro, with higher volumes of precipitation, mainly in the months of December, March and April. Scarton et al. (2023) evaluated the relationship between meteorological and geographic variables on the agronomic performance of soybeans. These authors revealed that altitude was the main variable that determined soybean grain productivity. The greater effect of altitude was also observed by Sarturi et al. (2022) in maize.
During development, for a potential of 6 ton ha-1, soybeans require approximately 800 mm of water, with regular distribution of rainfall
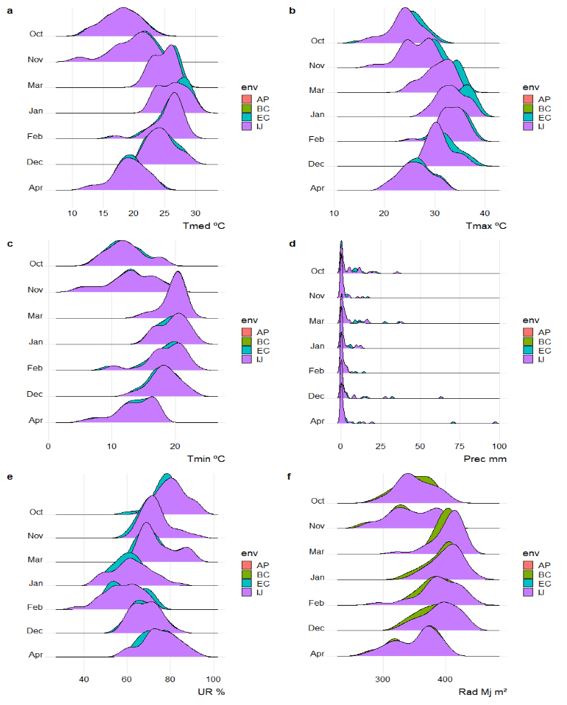
Fig 1. Mean air temperature, in ºC (a), maximum air temperature, in °C (b), minimum air temperature, in ºC (c), precipitation, in mm (d), relative air humidity, in % (e) and radiation global solar, in Mj m-2 (f), for different environments. AP: Augusto Pestana; BC: Boa Vista do Cadeado; EC: Eugênio de Castro; IJ: Ijuí.
being crucial, especially during the grain filling phase (Zanon et al., 2016), a period in which that the most pronounced water deficit occurred in the environments of Augusto Pestana and Boa Vista do Cadeado. A soybean plant has approximately 90% of its weight made up of water and this acts in essential physiological and biochemical processes (Taiz et al., 2017). Thus, the water deficit was a limiting factor for the development of soybeans, knowing the importance that water plays for the crop.
Genetic parameters
The variables number of seeds per plant (NSP) and grain weight per plant (SWP) presented high values of genotypic variance (Table 1), which indicates that the selection of genotypes based on the phenotype is efficient. To define the heritable portion of the genotypic variance, narrow-sense heritability values (h2) of 0.249 and 0.244 were estimated for NSP and SWP. The heritability found can be classified as medium according to Resende & Alves (2021), who defined that the magnitudes of individual heritabilities can be classified as low (0.01<h2<0.15), mean (0.15<h2<0.50) and high (h2>50). Heritability in the narrow sense indicates the amount of total observed variability that is due to additivity gene action (Singh; Singh; Singh, 2021). Carvalho et al. (2020), evaluating sources of variation in the genotype x environment interaction of soybean progenies conducted in different environments, found heritability values of 0.18, 0.12 and 0.16 for the variables number of grains per plant, thousand grain weight and grain productivity, respectively.
According to the classification by Resende & Duarte (2007), selection accuracy (Ac) results are considered: very high, Ac ≥ 0.90; high 70 ≤ Ac < 0.90; moderate, 0.50 ≤ Ac < 0.70; and low, Ac ≤ 0.50. In this case, the selection accuracy for the NSP and SWP variables, according to the previous classification, were considered very high for both variables (Table 1).
Selection for seed weight per plant
Of the total lines selected in the field for the variable seed weight per plant (SWP), 264 lines were obtained superior to the commercial controls (Figure 2), where 10.61%, 40.15%, 40.91% and 8 33% of the superior genotypes were selected in the environments of Augusto Pestana (AP), Boa Vista do Cadeado (BC), Eugênio de Castro (EC) and Ijuí (IJ), respectively. This demonstrates that progeny testing with joint analysis of experiments conducted in different environments is of great interest for breeding, as estimates of genetic parameters based on experiments conducted in a single environment are overestimated and may not result in expected selection gains.
The percentage of superior genotypes selected in different environments showed variation at the population level for the average seed weight per plant (Figure 2), where some populations had superior lines selected only in one environment. This was the case of the IRC 027 and IRC 038 populations, where 100% of the genotypes were selected in Augusto Pestana and Ijuí, respectively. The same behavior was observed for populations IRC 046, IRC 048 and IRC 050, with transgressive lineages selected only in Boa Vista do Cadeado. For the populations IRC 022, IRC 023, IRC 031, IRC 037, IRC 040, IRC 043, IRC 052, IRC 054 and IRC 057, selection of superior lineages was observed only in Eugênio de Castro, suggesting the existence of a specific interaction between population and environment.
Some transgressive genotypes were selected in two different environments, coming from the same base population and presenting a frequency equal to 50%, that is, with the same number of lineages per environment. This was verified for the populations IRC 001 and IRC 042, selected in Augusto Pestana and Eugênio de Castro, for the population IRC 005, with lineages selected in Augusto Pestana and Boa Vista do Cadeado and for the population IRC 026, with the same number of superior genotypes selected in Augusto Pestana and Ijuí. Thus, it can be inferred that environmental factors affect characters
Table 1. Estimates of genetic parameters for the variables number of seeds per plant (NSP) and seed weight per plant (SWP, g).
| REML | Variables | |
|---|---|---|
| NSP | SWP | |
| σ2G | 21081.400522 | 544.342594 |
| σ2E | 53.111983 | 13.113001 |
| σ2P | 21140.357133 | 557.951234 |
| H2 | 0.997211 | 0.975610 |
| h2 | 0.249300 | 0.243902 |
| Ac | 0.999547 | 0.996551 |
| OM | 34.478737 | 11.853111 |
σ2G: genotypic variance; σ2E: environmental variance; σ2P: phenotypic variance; H2: heritability in the broad sense; h2: heritability in the strict sense; Ac: genotype selection accuracy; OM: overall mean.
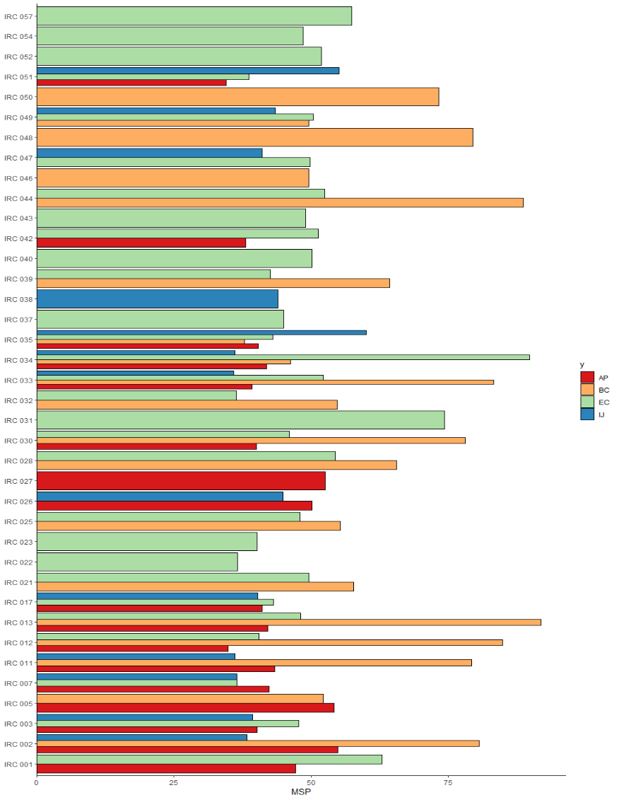
Fig 2. BLUP (Best Linear Unbiased Prediction) for the average seed weight per plant (MSP, g) of F2 lines grown in different environments. AP: Augusto Pestana (red); BC: Boa Vista do Cadeado (orange); EC: Eugênio de Castro (light greend); and IJ: Ijuí (blue).
through different stimuli and physiological mechanisms (Falconer, 1987), modifying phenotypic expression.
In the context of the selection of superior lineages in just two environments, genotypes were also identified in the environments of Boa Vista do Cadeado and Eugênio de Castro, based on massal selection within the IRC 021 populations (42.9 and 57.1%), IRC 025 (84.6 and 15.4%), IRC 028 (66.7 and 33.3%), IRC 032 (90 and 10%), IRC 039 (90.9 and 9.1%), IRC 044 (60 and 40%), with variation in the number of superior individuals obtained (Figure 2). It was also possible to select superior genotypes for the IRC 047 population in Eugênio de Castro and Ijuí, with percentages of 66.7 and 33.3%, respectively.
For the other populations, superior lines were selected in more than two locations, which suggests the existence of a broader interaction of these genotypes with the environments tested. Superior lineages were obtained in Augusto Pestana, Boa Vista do Cadeado and Ijuí for populations IRC 002 (14.3; 71.4 and 14.3%) and IRC 011 (11.1; 77.8 and 11.1%). Considering the segregation for IRC 003, IRC 007, IRC 017, and IRC 051, superior genotypes were obtained in the Augusto Pestana, Eugênio de Castro and Ijuí environments, with similar distribution for the number of individuals, with the exception of the IRC 017 population, in which 50% of the superior lineages were obtained in the Augusto Pestana environment. Transgressive genotypes were also
identified for the populations IRC 012 (16.7; 33.3 and 50%), IRC 013 (25; 37.5 and 37.5%) and IRC 030 (16.7; 16. 7 and 66.7%) in the environments of Augusto Pestana, Boa Vista do Cadeado and Eugênio de Castro, respectively, in addition to the IRC 049 population, where superior genotypes were obtained in the environments of Boa Vista do Cadeado, Eugênio de Castro and Ijuí.
Lineages transgressive to the average seed weight per plant were identified in four different environments (Figure 2) only for the populations IRC 033, IRC 034 and IRC 035. Considering the population IRC 033, 62.5% of the genotypes were selected in Boa View of Cadeado, with 12.5% for other environments. For IRC 034, selection percentages of 28.6 were observed for Augusto Pestana and 42.9% for Boa Vista do Cadeado, in addition to 14.3% for Eugênio de Castro and Ijuí. The largest number of superior lineages for the IRC 035 population was selected in Eugênio de Castro, totaling 63.6%, with 18.2% for Augusto Pestana and 9.1% for Boa Vista do Cadeado and Ijuí.
The genotypes that were selected demonstrate high potential for additive genetic gains, considering that the general average for BLUP was 11.85 g for the SWP variable (Table 1), knowing that all genotypes that were superior produced more than 100 seeds per plant (Figure 3). It can be considered that the midpoint of the BLUP for the average seed weight per plant (Figure 2) is at SWP = 50 grams, where some
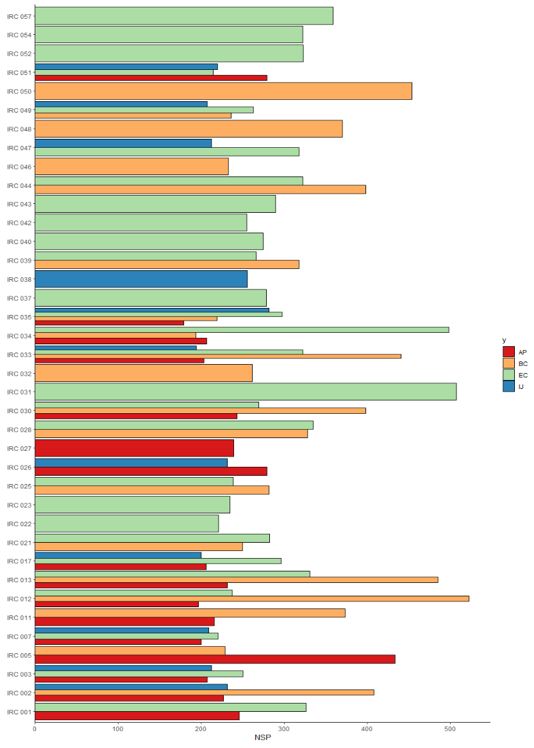
Fig 3. BLUP (Best Linear Unbiased Prediction) for the average number of seeds per plant (NSP) of F2 lines grown in different environments. AP: Augusto Pestana (red); BC: Boa Vista do Cadeado (orange); EC: Eugênio de Castro (light greend); and IJ: Ijuí (blue).
selected lines exceeded this mark for the variable, resulting in a high potential for gains for these individuals and deserve to be highlighted. This was the case with the superior lineages selected for populations IRC 002, IRC 005, IRC 012, IRC 013, IRC 031, IRC 034 and IRC 050, which demonstrated great potential. For lines transgressive to commercial controls, for the SWP variable, the values start from 34.28 g, reaching up to the line that produced 142.09 g, being higher than those found by Volpato (2016) evaluating the selection of F2 and F3 progenies of the soybean for grain production, where the predicted values for seed weight per plant varied between 29.17 and 31.63 g, considering only the genotypes that were superior.
Selection for number of seeds per plant
The BLUP for the average number of seeds per plant is shown in figure 3, where variation in the percentage of transgressive genotypes to commercial controls was also identified in relation to the base populations segregated in different environments. For Augusto Pestana, Boa Vista do Cadeado, Eugênio de Castro and Ijuí, there were percentages of 11.72%, 39.93%, 41.76% and 6.59% of a total of 273 superior lineages, considering all plants that were selected in the field. This shows that, in general, the genotypes expressed a certain affinity with higher altitude environments, even with similar climatic conditions during the crop cycle.
Of a total of 38 segregating populations conducted in different environments, 16 of them contributed superior lineages selected in just one environment (Figure 3). For the IRC 027 population, transgressive
genotypes were identified only in Augusto Pestana (AP). The same behavior was observed for IRC 038 in Ijuí (IJ), where all superior genotypes were selected for this population. In the same sense, 100% of the superior lineages for populations IRC 032, IRC 046, IRC 048 and IRC 050 were selected in the Boa Vista do Cadeado (BC) environment. For IRC 022, IRC 023, IRC 031, IRC 037, IRC 040, IRC 042, IRC 043, IRC 052, IRC 054 and IRC 057 all lines that demonstrated superiority were selected in Eugênio de Castro (EC).
In general, for the superior genotypes that were selected for each population, a similar percentage distribution was observed between the average seed weight per plant and the average number of seeds per plant (Figure 3), with changes only in the number of individuals on some occasions. This was the case of populations IRC 005 (33.3 and 66.7%) and IRC 011 (22.2 and 77.8%), with transgressive lineages selected only in Augusto Pestana and Boa Vista do Cadeado. In the same sense, superior genotypes were selected only in two locations for the populations IRC 025 (BC – 82.4%; EC – 17.6%), IRC 039 (BC – 81.8%; EC – 18.2%) and IRC 047 (EC – 70%; IJ – 30%).
Selection of transgressive lines in the number of seeds per plant was verified in more than three locations, where only some populations suffered changes in the percentage of genotypes for each environment (Figure 3). This behavior was observed for populations IRC 002 (33.3; 55.6 and 11.1%) and IRC 003 (28.6; 42.9 and 28.6%), which had superior lineages selected in Augusto Pestana, Boa Vista do Cadeado and Ijuí. Likewise, for the populations IRC 013 (33.3% for each environment), IRC 030 (20; 13.3 and 66.7%) and IRC 034 (33.3; 50 and 16.7%), with
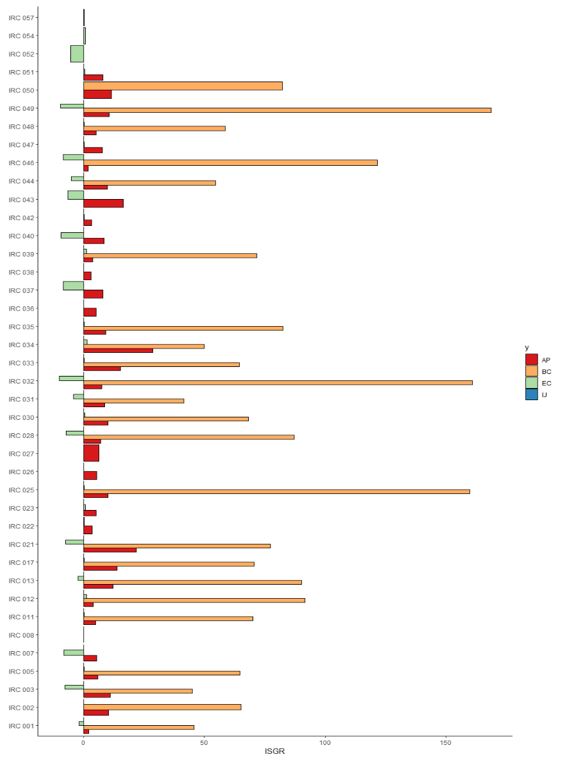
Fig 4. Index of Genetic Selection for Resilience (IGSR∆) of F2 lines conducted in different environments. AP: Augusto Pestana (red); BC: Boa Vista do Cadeado (orange); EC: Eugênio de Castro (light greend); and IJ: Ijuí (blue).
except for the Ijuí environment, superior genotypes were selected for the other three environments. Transgressive lineages were identified for IRC 017 and IRC 051, showing similar percentages per location, with the exception of the Boa Vista do Cadeado environment, where there were no superior genotypes for these populations.
Therefore, it can be seen that the superior lines that were selected also have a high potential for the number of seeds per plant (Figure 3), knowing that all genotypes exceeded the range of 100 seeds per plant. This is highly important for breeders, knowing that autogamous species, such as soybeans, increase the number of homozygous alleles with each generation of self-fertilization.
Index of genetic selection for resilience (IGSR∆)
The Index of Genetic Selection for Resilience (IGSR∆) makes a relationship between the variables of interest evaluated for each selected genotype, based on the values observed for the controls (Figure 4). The calculation considers real meteorological data for the cultivation cycle, making a comparison with air temperature and rainfall values considered optimal for the development of soybean crops. Therefore, the lower the IGSR∆ values, the more resilient the evaluated genotypes are.
The Boa Vista do Cadeado (BC) environment contributed to the selection of superior genotypes, transgressive to their respective parents, but proved to be a highly demanding environment for gene expression to occur in a way that results in considerable productivity, as it presents indices with high values, mainly for populations IRC 025, IRC 039 and IRC 042, which presented IGSR∆ greater than 150. In the Eugênio de Castro (EC) environment, the selected lines showed
greater resilience, since they presented the lowest IGSR∆. Thus, it can be inferred that the lines selected in Eugênio de Castro have high potential for the development of soybean cultivars with high resilience. The environments of Ijuí (IJ) and Augusto Pestana (AP) also showed superior selection gains for resilience compared to the BC environment, with reduced IGSR∆ values, with emphasis on the IRC 001, IRC 008 and IRC 046 populations.
Materials and Methods
Characterization of germplasm and environments
Thirty eight soybean F2 segregating populations were evaluated in four environments (Supplementary Figure 1) in the state of Rio Grande do Sul, Brazil, namely: Augusto Pestana (28º 25’ 58” S, 54 º00’ 20’’ W and 283 m altitude); Boa Vista do Cadeado (28º31’ 19’’ S and 53º 46’ 16’’ W and 428 m altitude); Eugênio de Castro (28º31’ 02’’ S, 54º06’ 19” W and 345 m altitude); Ijuí (28º 18’ 25’’ S, 54º 01’ 51’’ W and 299 m altitude).
The soil in the four environments is characterized as a Typical Distroferric Red Oxisol (U. M. Santo Ângelo), with a deep, well-drained profile, dark red color and high clay content. According to the Köeppen climate classification, all locations have a climate that falls into type Cfa, which consists of a subtropical climate with hot summers, where temperatures are above 22 ºC in summer and with more than 300 mm of rain in the driest month.
The segregating populations of the F2 generation of soybeans come from the UNIJUÍ Grain Genetic Improvement Program. The F1 generation seeds were obtained from crossing, in a greenhouse, between the parents described in supplementary table 1. A self-fertilization cycle of the F1 plants was carried out to obtain the population of F2 plants.
Experimental design and data collection procedure
To evaluate the populations, an incomplete block design augmented with interspersed controls was used. The experiments in Augusto Pestana, Boa Vista do Cadeado, Eugênio de Castro and Ijuí were sown on the dates 11/09/2022, 10/19/2022, 11/18/2022 and 12/22/2022, respectively. In the four environments, sowing was carried out with the aid of a seeder-fertilizer, with a sowing density of 18 seeds per linear meter. All management was carried out preventively, in order to avoid the occurrence of biotic effects on the plants. At full physiological maturity, the best performing plants from each population were selected using the massal selection method stratified by environments. The seeds from the selected plants were sent to a forced air greenhouse to correct humidity to 13%. The variables number of seeds per plant (NSP) and seed weight per plant (SWP, g) were determined. The number of seeds per plant was determined using the RStudio software (R Core Team, 2023), using the pliman (Plant Image Analysis) data analysis package (Olivoto, 2022).
Statistical analysis
The variance components and genetic parameters were determined using the REML/BLUP procedure (restricted maximum likelihood/best unbiased linear prediction), using the methodology proposed by Resende (2007). The statistical model proposed by the author is y = Xf + Zg +Wb + e, where y is the data vector, f is the vector of effects assumed to be fixed (average of the population of main treatments), g is the vector of effects genotypic (assumed to be random), and is the vector of errors or residues (random), with capital letters representing the incidence matrices for the aforementioned effects.
To determine the superiority of the plants that were selected for the variables number of seeds per plant and seed weight per plant, 25 commercial soybean controls were used (M 6100 I2X, TEC 6702 IPRO, ST 6111 I2X, TMG 2751 IPRO, TMG 3621 IPRO, FT 4464 I2X, BRS 2553 158 RR, FT 1060 I2X, FT 4462 I2X, BRS 2558 XTD, ST 580 I2X, ST 490 I2X, ST 611 I2X), which were conducted in the same environment as Augusto Pestana.
As a way to determine the genetic gains of each population, the Index of Genetic Selection for Resilience (IGSR∆) was calculated using the following equation:
\(IGSR\mathrm{\Delta} = \left\lbrack \left( \frac{NSC - NSL}{SNS} \right)x\left( \frac{SWC - SWL}{SSW} \right)x\left( \frac{PRECI - PRECR}{SPREC} \right)x\left( \frac{TMEANI - TMEANR}{STMEAN} \right) \right\rbrack\), where:
NSC: prediction of the average number of seeds of pure line controls; NSL: prediction of the number of seeds of the lines; SSW: sample standard deviation of the prediction of the variable NSP; SWC: prediction of the average seed weight of pure line controls; SWL: prediction of the seed weight of the lines; SSW: sample standard deviation of the prediction of the variable SWP; PRECI: ideal precipitation for the crop, where the value of 720 mm was used; PRECR: real precipitation; SPREC: sample standard deviation of actual precipitation; TMEANI: ideal mean air temperature for the culture, where the value of 25ºC was used; TMEANR: average real air temperature; STMEAN: sample standard deviation of the variable. All analyzes were performed using the R software (R Core Team, 2023).
Conclusion
The progenies selected from populations IRC 002, IRC 005, IRC 012, IRC 013, IRC 031, IRC 034 and IRC 050 have a high potential for number of seeds per plant. The progenies selected from populations IRC 002, IRC 011, IRC 012, IRC 013, IRC 033, IRC 034 and IRC 044 have a high potential for seed weight per plant. The progenies selected from populations IRC 032 and IRC 049 showed the greatest resilience to abiotic stresses.
References
Albuquerque JRT, Lins HA, Santos MG, Freitas MAM, Oliveira FS, Souza ARE, Silveira LM, Nunes GHS, Barros Júnior AP, Vieira PFMJ (2022) Adaptability and Stability of soybean (Glycine max L.) genotypes in semiarid conditions. Euphytica. 218(61).
Almeida HC, Soares APL, Poersch NL, Izidorio TH, Teodoro LPR, Teodoro PE (2021) Adaptability and stability of soybean cultivars in the region of Chapadões. Rev Ceres. 68(4).
Carvalho IR, Silva JAG, Ferreira LL, Szareski VJ, Demari GH, Facchinello PHK, Moura NB, Schneider RO, Rosa TC, Magano DA, Souza VQ (2020) Relative contribution of expected sum of squares values for soybean genotypes × growing environments interaction. Austr J Crop Sci. 14(3).
Chen Y, Xiong Y, Hong H, Li G, Gao J, Guo Q, Sun R, Ren H, Zhang F, Wang J, Song J, Qiu L (2023) Genetic dissection of and genomic selection for seed weight, pod length and pod width in soybean. The Cr Journ. 11(3).
Facchinello PHK, Carvalho IR, Streck EA, Aguiar GA, Goveia J, Feijó M, Pereira RR, Oliveira VF, Fagundes PRR, Maia LC, Lautenchleger F, Magalhães Júnior AM (2021) Predicted genetic gains weighted by selection pressures for grain quality in irrigated rice. Pesq Agr Bras. 56(e02497).
Falconer DS (1987) Introduction to quantitative genetics. Viçosa:UFV.
Laviola BG, Silva DAS, Juhász ACP, Rocha RB, De Oliveira RJP, Albrecht JC, Alves AA, Rosado TB (2014) Agronomic performance and genetic gain by Jatropha selection in three regions of Brazil. Pesq Agrop Bras. 49(5).
Loro MV, Carvalho IR, Silva JAG, Moura NB, Hutra DJ, Lautenchleger F (2021) Artificial intelligence and multiple models applied to phytosanitary and nutritional aspects that interfere in the physiological potential of soybean seeds. Braz J Agric. 96.
Loro MV, Carvalho IR, Sfalcin IC, Pradebon LC (2022) Decomposition of white oat phenotypic variability by environmental covariates (2022) Braz J Agric. 97(3).
Olivoto T (2022) Pliman: an R package for plant and image analysis. Met Ecol Evol. 13(1).
Pradebon LC, Carvalho IR, Loro MV, Port ED, Bonfada B, Sflacin IC, Santos IJ, Bruinsma GMW, Silva JAG, Alban AA, Challiol MA (2023) Soybean adaptability and stability analyzes to the organic system trough AMMI, GGE Biplot and mixed models methodologies. Ciênc Rur. 53(9).
R Core Team (2023) R: a language and environment for statistical computing. Vienna, Austria.
Resende MDV (2007). SELEGEN-REML/BLUP: statistical system and computerized genetic selection via linear mixed models. Colombo, Embrapa Forests.
Resende MDV, Alves RS (2021) Genética: estratégias de melhoramento e métodos de seleção. Embrapa.
Resende MDV, Duarte JB (2007) Precisão e controle de qualidade em experimentos de avaliação de cultivares. Pesq Agrp Trop. 37(3).
Sangiovo JP, Carvalho IR, Pradebon LC, Loro MV, Port ED, Scarton VDB, Zuse GH, Sfalcin IC, Foleto EE (2023) Selection of soybean lines based on a nutraceutical ideotype. Neotr Agric Journ. 10(3).
Said AA, Macqueen AH, Shawky H, Reynolds M, Juenger TE, El-Soda M (2022) Genome-wide association mapping of genotype-environment interactions affecting yield-related traits of spring wheat grown in three watering regimes. Envir Exp Bot. 194(104740).
Sarturi MVR, Texeira CAMB, Carvalho IR, Demari GH, Loro MV, Pradebon LC, Port ED (2022) Prediction of corn grain productivity as a function of altitude and plant population. Rev Agric Neotr. 9(4).
Scarton VDB, Carvalho IR, Pradebon LC, Loro MV, Alban AA, Challiol MA, Sausen NH, Braga PMF, Sflacin IC (2023) Influence of meteorological variables and geographic factors in the selection of soybean lines. Rev Agric Neotr. 10(3).
Singh DP, Singh AK, Singh A (2021) Plant breeding and cultivar development. Oxford, Academic Press.
Taiz L, Zeiger E, Moller I, Murphy A (2017) Plant physiology and development. Porto Alegre, Artemd, 6ed.
USDA – United States Departament of Agriculture (2023) Word agricultural production – circular series, may 2023. Available in:<https://apps.fas.usda.gov/psdonline/circulars/production.pdf.>
Zanon AJ, Streck NA, Grassini P (2016) Climate and management factors influence soybean yield potential in a subtropical environment. Agron Journ. 108(4).