ISSN:1835-2707
Aust J Crop Sci. 18(08):466-470 (2024)
https://doi.org/10.21475/ajcs.24.18.08.pne49
Research Note
Seed size improves vigor and emergence of Cenostigma tocantinum seedlings
Ruan Santos de Farias1, Sérgio Heitor Sousa Felipe2, Vanessa Nascimento Brito1*
1Universidade Federal do Pará, Altamira, Brazil
2Universidade Estadual do Maranhão, São Luís, Brazil
Abstract
Cenostigma tocantinum is a tree native to Brazil, with high timber and urban landscape potential. This species is propagated by seeds; however, few studies have reported the production of seedlings with higher quality standards. This study aimed to evaluate the emergence and initial growth of C. tocantinum seedlings of contrasting seed sizes (small and large). The design was completely randomized with two treatments [small seed (dimensions of length 1.20 cm; width 1.00 cm; thickness 0.32 cm; and fresh weight 0.18 g) and large seed (dimensions of length 1.52 cm; width 1.35 cm; thickness 0.39 cm; and fresh weight 0.37 g)] and four replicates of 25 seeds each. One hundred seeds from each group were randomly selected for measurements of the length, thickness, width and fresh weight. During the experiment, the following emergence and emergence speed index (ESI) were assessed. After 10 days of sowing, the shoot and root lengths, stem diameter, and shoot and root dry weights were assessed. The results showed that the length, width, thickness, and fresh weight of large seeds were 27, 35, 22, and 106% greater, respectively, than those of the small seeds. Regarding emergence (ranging from 21.8 to 23.8%) and ESI (ranging from 4.4 to 4.8), no significant differences were observed (P > 0.05) between large and small seed sets. More importantly, large seeds favored the development of seedlings with shoot length, root length, collar diameter, shoot dry weight, and root dry weight that were 29, 38, 17, 93, and 109% higher, respectively, than those of small seeds. Thus, C. tocantinum seeds can be stratified into two different sizes (small and large), with large seeds providing the initial growth of more vigorous seedlings (e.g., growth, development, and dry weight). The findings of this study can aid seedling producers in improving their product to meet higher quality standards.
Keywords: emergence; forest seed; seedling production; size classes; vigor.
Introduction
Cenostigma tocantinum (Fabaceae family) is a tree species native to Brazil, with a wide occurrence in the Amazon Biome, especially in the Terra Firme Forest and Várzea Forest vegetation (Gaem, 2023). This tree is of great significance for its high timber value; however, it is widely used in urban landscaping projects (Almeida et al., 2015; Gaem, 2023). C. tocantinum is a commonly used native species in the afforestation of cities in the North of Brazil owing to its desirable characteristics, such as straight trunk, fast growth, slightly aggressive root system and leafy canopy, which in turn provides efficient shading without releasing a large number of leaves (Lima Junior et al., 2022).
C. tocantinum seedlings are primarily produced from seeds (Lima Junior et al., 2022). However, to achieve successful forest restoration in habitats with a high degree of degradation, strategies that guarantee seedling production with greater vigor should be developed (Guariz et al., 2021). Thus, the physical (e.g., size, weight, color, and moisture content) and physiological (e.g., germination and vigor) characteristics of seeds are essential for the production of seedlings with a higher quality standard (Leão et al., 2018a; Leão et al., 2023).
In agricultural crops, genes affecting seed size are well reported. This can be exemplified by the size and shape of rice seeds in what is widely accepted to be complex traits controlled by polygenes (GS3, GW2, qSW5/GW5, and GIF1) and numerous quantitative trait loci (Yan et al., 2011). In parallel, seed reserves have important effects on initial seedling growth, improving growth and development (Kennedy et al., 2004). Furthermore, the environment also plays a role in this entire process, since the different plant species require specific requirements regarding nutrition, water availability, photoperiod, and others (Fenner, 1992). In contrast, native trees from the Brazilian Amazon have little or no information available in scientific literature.
Seed size is a characteristic that presents great variability and is defined by length, width, and thickness (Leão et al., 2018b). For numerous native tree species, seed size can influence germination potential, because smaller seeds generally have lower reserves, which can affect their ability to germinate and grow, particularly in dry environments with low soil fertility (Kennedy et al., 2004; Ambika et al., 2014). In contrast, larger seeds tend to produce more vigorous and resistant seedlings, with a greater capacity for survival and growth under adverse environmental conditions (Mota et al., 2019).
In this study, we investigated the germination and emergence of C. tocantinum seedlings with contrasting seed sizes (small and large) to generate information for seedling production with greater quality standards for this species.
Results and discussion
For native tree species of high commercial value that have not been studied in plant genetic improvement programs, developing feasible strategies is essential for seedling producers to obtain seedlings with higher quality standards (Leão et al., 2018b). Manual stratification of C. tocantinum seeds into two groups, small and large, showed that large seeds tend to produce seedlings with greater vigor and dry mass.
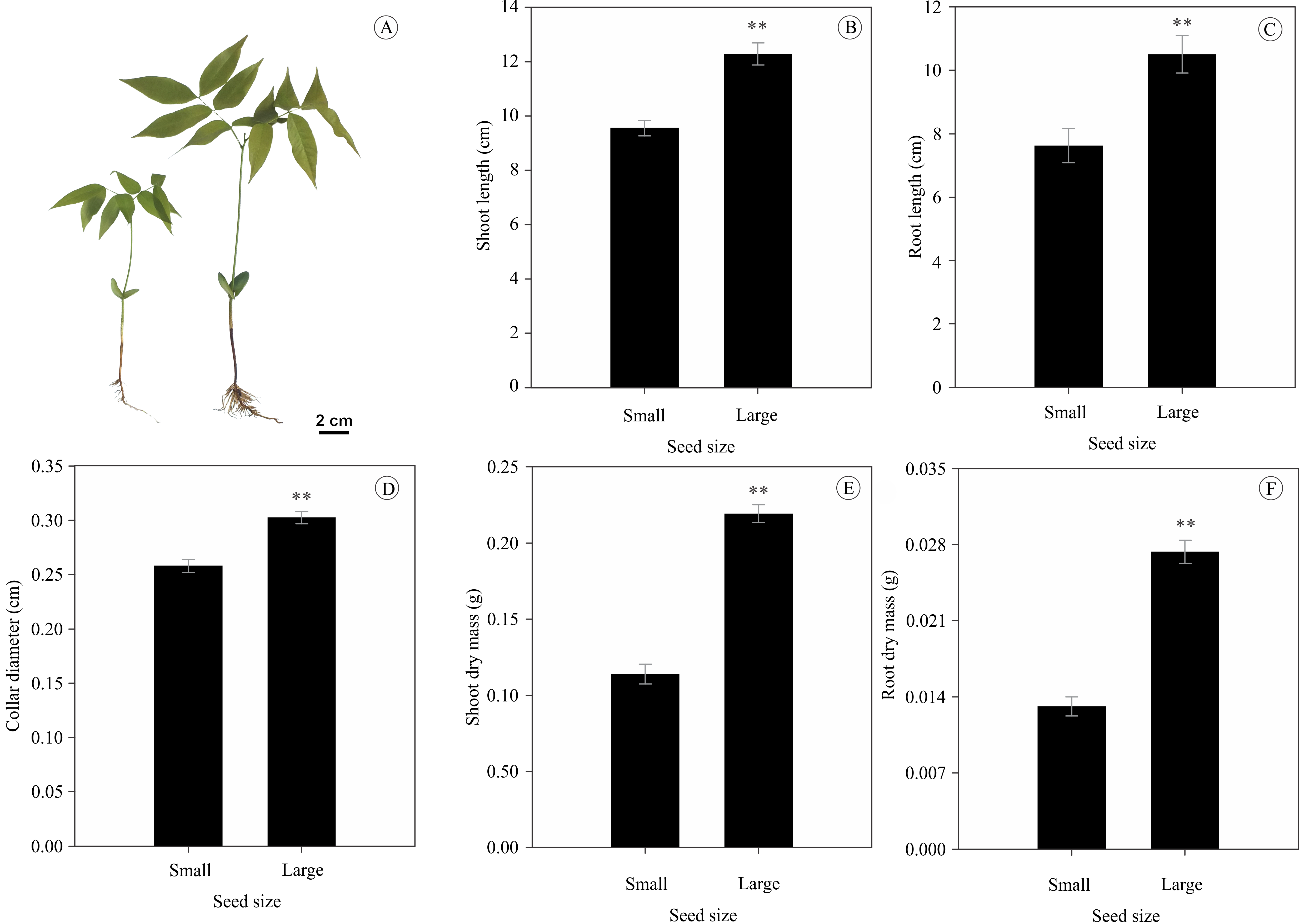
C. tocantinum seeds showed significant differences in all the evaluated size and fresh weight traits (P ≤ 0.01). Notably, the large seeds had a length, width, thickness, and fresh weight up to 27, 35, 22, and 106% greater, respectively, than those in the small seeds (Figure 2).
Here, the two distinct groups of seed sizes (small and large) in this study indicate that the presence of phenotypic variability can be exploited by seedling producers, as it is possible to stratify seeds into these groups to obtain better results in seedling production. This result can be explained by genetic and environmental factors (climate and soil), in line with the fact that C. tocantinum is a non-domesticated species, which in turn leads to the production of fruits and seeds with variable phenotypic aspects of size and weight (Leão et al., 2018b). It should be noted that there are no conclusive studies on the reproductive system of C. tocantinum, and allogamy is suspected (Almeida, 2014). Based on this, and considering the fact that the mother trees of C. tocantinum in the current study were in an environment without pollen flow control, it is difficult to infer how much this may have affected seed size.
We emphasize that although genetic and environmental factors influence seed traits, we cannot neglect that the formation of smaller seeds is also due to positional issues with fruits. In general, a fruit can contain up to six seeds distributed in a dry legume type with a narrower base than the apex; the seeds formed in this region of the base during morphogenesis tend to be smaller than those formed in the center or apex (Lima Júnior et al., 2022).
The percentage of seedling emergence for small and large seeds was 21.8 and 23.8%, respectively. Although large seeds provided superior emergence compared with that in small seeds, the difference was not statistically significant (P > 0.05). Furthermore, the average germination speed index tended to be higher for small seeds than for large seeds (ranging from 4.4 to 4.8), but with no statistical difference (P > 0.05). These results clearly show that seedling emergence and the speed at which it occurs are independent of the seed size. In general, seed size does not always interfere with emergence (Cazetta et al., 1995), which corroborates the results of the present study.
Although seed size can be indicative of physiological quality (Popinigis, 1985), seedling emergence was not significantly different between the two sizes of C. tocantinum seeds, as both smaller and larger seeds were collected directly from the trees during the same period. This finding indicates that physiological differences within the same seed lot may be associated with other factors, such as a lack of standardization during harvest (Leão et al., 2015; Krzyzanowski et al., 2022).
Small and large seeds showed significant differences in growth characteristics and seedling dry mass (P < 0.01). Large seeds favored the development of seedlings with shoot length, root length, collar diameter, shoot dry mass, and root dry mass being up to 29, 38, 17, 93, and 109% higher, respectively, than those in small seeds (Figure 3).
The response patterns in seedling growth and initial development clearly showed a dependence on seed size. This response may be associated with the formation phase of the fruits and seeds, which received the greatest amount of assimilates during their development; well-formed embryos had greater amounts of reserves at the end of growth and development (Dresch et al., 2013). These substances are mobilized during embryo growth, where during seedling development their degradation products are used as a source of energy and substrate for cellular structures. Thus, later in the germination phase, the development of more vigorous seedlings with greater biomass will potentially occur considering that the entire endosperm is absorbed by the embryo to resume growth and seedling formation (Mota et al., 2019; Doll and Ingram, 2022).
Our findings offer possibilities to explore seed size in C. tocantinum as a strategy for obtaining seedlings with higher quality standards. In future work, it will be important to address seed size by comparison different mother trees to understand the impact of genotype on seed size of this species.
Materials and Methods
Plant material and seed extraction
C. tocantinum fruits were harvested directly from the crown of three mother trees located in Altamira, Pará, Brazil. The trees were selected with a minimum distance of 50 m from each other, in order to avoid the selection of half-sibs. Furthermore, the absence of phytosanitary problems (attack by insect pests and diseases) was considered. After harvesting, the fruits were dried in a shaded and protected environment, followed by manual extraction and seed lot formation as described by Leão et al. (2015). Then, a seed lot was formed containing seeds from three mother trees. The experiment was conducted at the Microbiology Laboratory of the Federal University of Pará, Altamira Campus, Pará, Brazil.
Contrasting seed sizes
The seed lot was subdivided into two groups of seeds of different sizes: small and large (Table 1; Figure 1). Subdivisions were performed manually, considering the size of the seeds. One hundred seeds from each group were randomly selected for size measurements using a digital caliper with a precision of 0.01 mm. The length was measured from the apex (region of the hilum) to the base (region opposite the hilum), and thickness and width were measured in the intermediate region. In addition, fresh weight was measured using a precision analytical balance.
Seedling emergence
The emergence test was conducted under ambient conditions [natural light conditions, relative humidity air (∼ 80%), and a temperature of approximately 27 ± 3 °C], consisting of four replicates of 25 seeds, on sand substrate (previously washed in running water and sterilized in an oven at 120 °C for 24 h) with sawdust (sterilized in an oven at 120 °C for 4 h) in a ratio of 1:1. They were then packed in plastic trays. Irrigation was performed daily whenever necessary (at low substrate humidity).
During the experiment, the following physiological traits were assessed: i) emergence (E;%): the percentage of seeds that emerged was calculated using the following equation: $E = \left( \sum_{i = 1}^{k}\frac{n_{i}}{n} \right)\ 100$; where i = 1 is the first day of evaluation of the emergence test, k is the last day of evaluation of the emergence test, ni is the number of emerged seeds on day i, and n is the total number of seeds placed; and ii) emergence speed index (ESI): the ESI was calculated using the following equation (Maguire, 1962): $ESI = \ \sum_{}^{}\left( \frac{n_{i}}{t_{i}} \right)$; where ni is the number of seeds emerging at time i, and ti is the time after test installation.
After 10 days of sowing, the shoot and root lengths, stem diameter, and shoot and root dry weights were assessed.
Experimental design and statistical analysis
The design was completely randomized with two treatments (small and large seed sizes) and four replicates of 25 seeds. Seed data (length, width, thickness, and fresh weight) were evaluated using descriptive statistics by box plot, and seedling data (percentage of emergence; shoot and root length, stem diameter, and dry weight of shoot and root) were compared using a two-tailed t-test (P ≤ 0.05 and P ≤ 0.01).
Conclusion
C. tocantinum seeds can be stratified into two different sizes (small and large). Notably, large seeds lead to the initial growth of more vigorous seedlings (greater growth, development, and dry mass). The results of this study can be applied to obtain vigorous and resistant seedlings with greater potential to survive under adverse environmental conditions.
Acknowledgements
The authors thank the Universidade Federal do Pará (UFPA) for the Scientific Initiation Scholarship Program for the first author [grants Edital 04/2021; Pró-Reitoria de Pesquisa e Pós-Graduação].
References
Almeida FV (2014) Diversidade genética entre e dentro de populações de Cenostigma tocantinum Ducke. Orientadora: Maria Teresa Gomes Lopes. Masters’s Thesis, Universidade Federal do Amazonas, Manaus, Brazil. 42p. Available online: https://tede.ufam.edu.br/handle/tede/4266
Almeida FVD, Lopes MTG, Valente MSF, Bentes JLDS (2015) Diversidade genética entre e dentro de populações de Cenostigma tocantinum Ducke. Sci. For. 43(108): 753-762. https://www.ipef.br/publicacoes/scientia/nr108/cap01.pdf
Ambika S, Manonmani V, Somasundaram G (2014) Review on effect of seed size on seedling vigour and seed yield. Res J Seed Sci. 7(2): 31-38. https://doi.org/10.3923/rjss.2014.31.38
Cazetta JO, Sader R, Ikeda M (1995) Efeito do tamanho no desempenho germinativo de sementes de feijoeiro (Phaseolus vulgaris L.). Científica. 23(1):65-71.
Doll NM, Ingram GC (2022) Embryo–endosperm interactions. Annual Review of Plant Biology.73: 293-321. https://doi.org/10.1146/annurev-arplant-102820-091838
Dresch DM, Scalon SDPQ, Masetto TE, Vieira MDC (2013) Germinação e vigor de sementes de gabiroba em função do tamanho do fruto e semente. Pesqui. Agropecu. Trop. 43(3): 262-271. https://doi.org/10.1590/S1983-40632013000300006
Fenner M (1992) Environmental influences on seed size and composition. Hortic. Rev. 13: 183-213.
Gaem PH (2020) Cenostigma in Flora e Funga do Brasil. Jardim Botânico do Rio de Janeiro. Availability in: <https://floradobrasil.jbrj.gov.br/FB78636>. Access on November 15, 2023.
Guariz HR, Oliveira HCD, Sperandio HV, Paula JCBD, Shimizu GD, Ribeiro Junior WA (2021) Germination potential of Hymenaea courbaril L. in different maturation stages. Semina Ciênc Agrar. 42(6, supl. 2): 3667-3684. https://doi.org/10.5433/1679-0359.2021v42n6SUPL2p3667
Kennedy PG, Hausmann NJ, Wenk EH, Dawson TE (2004) The importance of seed reserves for seedling performance: an integrated approach using morphological, physiological, and stable isotope techniques. Oecologia. 141: 547-554. https://doi.org/10.1007/s00442-004-1686-0
Krzyzanowski FC, Dias DCFS, França-Neto JB (2022) Deterioração e vigor da semente. Brasília, DF, Brazil: Embrapa Soja, 19 p. ilust. (Circular Técnica, 191). https://ainfo.cnptia.embrapa.br/digital/bitstream/doc/1151118/1/Circ-Tec-191.pdf
Leão NVM, Felipe SHS, Silva CE, Moraes ACS, Shimizu ESC, Gallo R, Freitas ADD, Kato OR (2018b) Morphometric diversity between fruits and seeds of mahogany trees (Swietenia macrophylla King.) from Parakanã Indigenous Land, Pará State, Brazil. Aust. J. Crop Sci. 12(3): 435-443. https://doi.org/10.21475/ajcs.18.12.03.pne879
Leão NVM, Felipe SHS, Gallo R, Shimizu ESC (2023) Genetic variability of Tachigali vulgaris trees based on seed morphophysiological traits. South. For. 85(3):1-9. https://doi.org/10.2989/20702620.2023.2226335
Leão NVM, Ohashi ST, Freitas ADD, Nascimento MRSM, Shimizu ESC, Reis ARS, Galvão Filho AF, Souza D (2011) Colheita de Sementes e Produção de mudas de espécies florestais nativas. Belém, PA, Brazil: Embrapa Amazônia Oriental, 47 p. ilust. (Documentos, 374). https://ainfo.cnptia.embrapa.br/digital/bitstream/item/124762/1/DOC-374-2Ed-ONLINE.pdf
Leão NVM, Shimizu ESC, Felipe SHS, Benchimol RL, Nascimento MRSM (2018a) Morfometria, germinação e sanidade de sementes de tachi-peludo. Encic Biosf. 15(27): https://doi.org/142-154. 10.18677/EnciBio_2018A60
Lima Junior MJV, Mendes MAS, Arruda YMBC (2022) Pau-pretinho: Cenostigma tocantinum Ducke. Inf. Abrates. 29(1-3): 23-25.
Maguire JD (1962) Speed of germination aid in selection and evaluation for seedling emergence and vigor. Crop Sci. 2(2): 176-177. https://doi.org/10.2135/cropsci1962.0011183X000200020033x
Mota NM, Fortini EA, Luz GR, Veloso MDDM, Fernandes GW, Nunes YRF (2019) Influência do tamanho e da escarificação dos diásporos na emergência e estabelecimento de Pterodon emarginatus. Pesqui. Florest. Bras. 39: 1-11. https://doi.org/10.4336/2019.pfb.39e201801743
Popinigis F (1985) Fisiologia da semente. Brasília, DF: AGIPLAN. 289 p.
Yan S, Zou G, Li S, Wang H, Liu H, Zhai G, Guo P, Song H, Yan C, Tao Y (2011) Seed size is determined by the combinations of the genes controlling different seed characteristics in rice. Theor. Appl. Genet. 123: 1173-1181. https://doi.org/10.1007/s00122-011-1657-x
Table
Table 1. Size classification of C. tocantinum seeds according to length, width, thickness, and fresh weight.
| Size | Length (cm) | Width (cm) | Thickness (cm) | Fresh weight (g) |
|---|---|---|---|---|
| Small | 1.20 ± 0.12 | 1.00 ± 0.13 | 0.32 ± 0.06 | 0.18 ± 0.06 |
| Large | 1.52 ± 0.07 | 1.35 ± 0.08 | 0.39 ± 0.06 | 0.37 ± 0.05 |
Figures
Figure 1. Cenostigma tocantinum seeds. Top row comprises small seeds, and bottom row comprises large seeds. The arrow indicates the hilum.
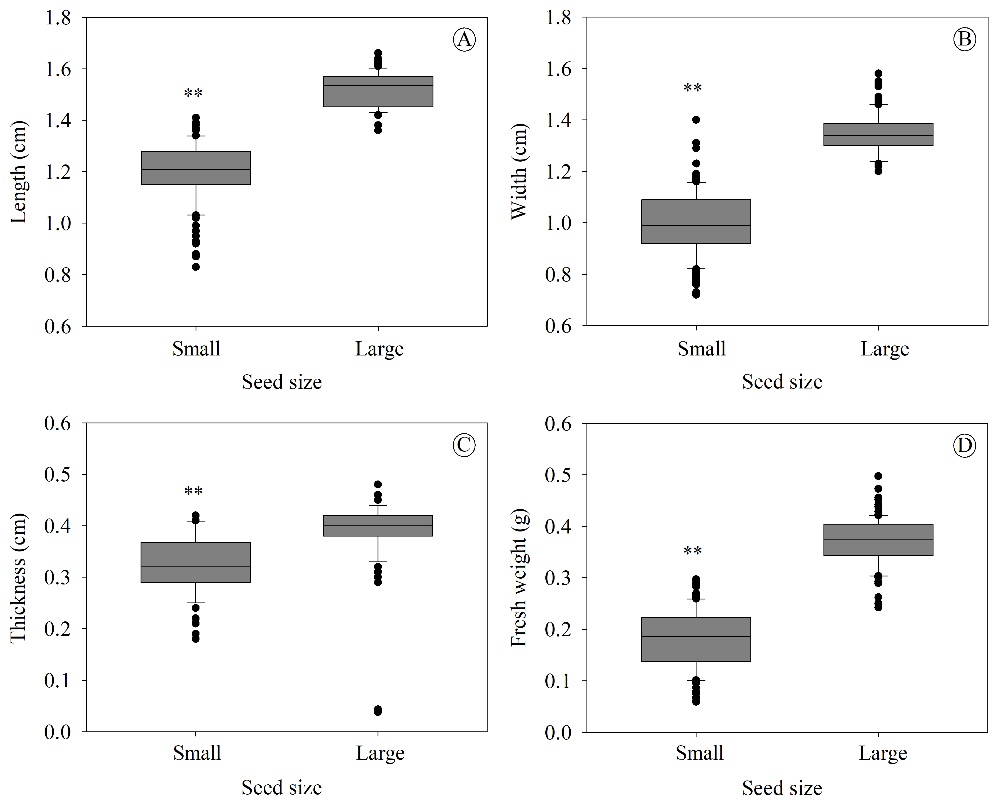
Figure 2. Box-plot of biometric traits of Cenostigma tocantinum seeds stratified into small and large seeds: length (A), width (B), thickness (C), and fresh weight (D) (n = 100). Asterisks indicate significant differences between treatments (*P ≤ 0.05 and **P ≤ 0.01, two-tailed t-test) and NS, non-significant.
Figure 3. Growth of Cenostigma tocantinum seeds based on size: seedlings from small (left) and large (right) seeds (A), shoot length (B), root length (C), collar diameter (D), shoot dry mass (E), and root dry mass (F) (n = 25). Asterisks indicate significant differences between treatments (*P ≤ 0.05 and **P ≤ 0.01, two-tailed t-test) and NS, non-significant.